Семенное размножение
Семена как органы размножения и расселения присущи только семенным растениям, как голо-, так и покрытосеменным. У споровых растений эту функцию выполняют споры. Семена образуются на материнском растении в огромном количестве, опадают с него и дают начало новым особям, подобным материнским. Чередование поколений при этом существует, но настолько замаскировано, что внешне совершенно не проявляется.
Важным отличием семенных растений от высших споровых является то, что гаметофиты, особенно женский, утратили способность к самостоятельному существованию и живут только за счет спорофита.
Семенные растения сделали огромный шаг по пути эволюции, освободившись от необходимости осуществлять половой процесс при помощи воды и адаптировавшись к наземной жизни. Это дало преимущества семенным растениям перед споровыми. Им не нужна вода для передвижения гамет, у них возник процесс опыления — перенос мужских гаметофитов по воздуху. По своей сущности семенные растения являются разноспоровыми, у них на спорофите формируются микро- и мегаспорангии с мейоспорами, возникающими в результате мейоза.
В отличие от споровых микро- и мегаспоры из спорангиев у семенных растений наружу не высыпаются, а прорастают внутри спорангиев под защитой их стенок и других защитных образований на материнском спорофите, т. е. развитие мужских и женских гаметофитов происходит внутри микро- и мегаспорангиев.
Семя формируется из семязачатка (семяпочки), представляющего собой видоизмененный мегаспорангии, внутри которого развивается женский гаметофит и происходит оплодотворение, после чего из зиготы появляется зародыш нового спорофита. Внешние покровы мегаспорангия превращаются в семенную кожуру, а семязачаток в целом превращается в семя. Формирование запасающей ткани в семенах голо- и покрытосеменных происходит по-разному.
Циклы развития и семенное размножение голосеменных
Циклы развития голосеменных довольно сходны, но различаются временем протекания отдельных этапов. Классическим примером для анализа этого процесса давно стала сосна обыкновенная (рис. 241). Голосеменные, как правило, — крупные деревья, особенно виды класса хвойных, представляют собой спорофит. На спорофите развиваются мужские и женские шишки. Мужские шишки после выхода пыльцы в окружающую среду засыхают и опадают. В женских шишках происходит формирование семян. Шишки — органы спороношения (совокупность мега- и микроспорофиллов, т. е. по своей природе являются органами бесполого размножения).
Женская шишка состоит из укороченной оси, с расположенными на ней кроющими пленчатыми чешуями. В пазухах последних развиваются утолщенные, впоследствии одревесневающие семенные чешуи, на верхней стороне которых расположены два семязачатка.
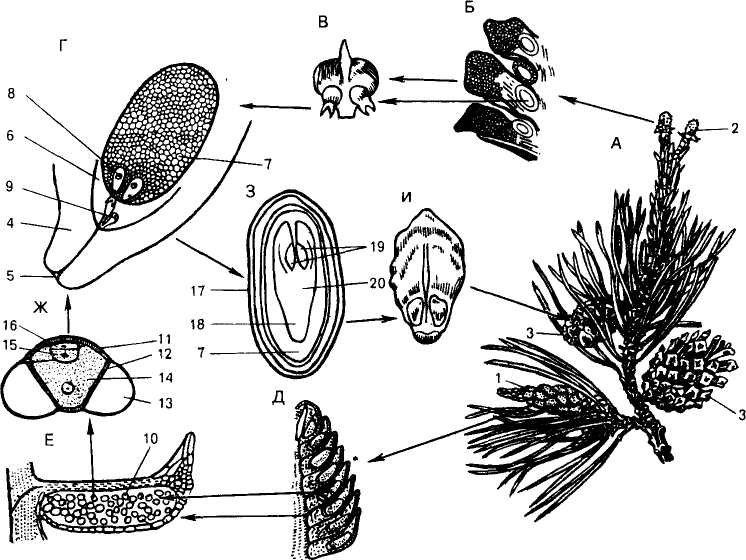
Рис. 241. Сосна обыкновенная (Pinus sylvestris L.):
А — ветка с шишками; Б — часть продольного разреза молодой женской шишки; В — семенная чешуя с семязачатками; Г — семязачаток в продольном разрезе; Д — часть продольного разреза мужской шишки; Е — продольный разрез микроспорофилла; Ж — пыльцевое зерно; 3 — продольный разрез семени; И — семенная чешуя зрелой шишки; 1 — группа мужских шишек; 2 — молодая женская шишка; 3 — зрелые женские шишки; 4 — интегумент, 5 — микропиле; 6 — нуделлус; 7 — эндосперм (женский гаметофят); 8 — архегоний; 9 — пыльцевая трубка со спермиями; 10 — микроспорангий; 11 — экзина; 12 — интина 13 — воздушный мешок; 14 -вегетативная клетка; 15 — антеридиальная клетка; 16 — проталлиальные клетки; 17 — кожура семени; 18 — зародышевый корешок; 19 — семядоли; 20 — гипокотиль
Семязачаток возникает в виде меристематического бугорка на молодой семенной чешуе. Из основания семязачатка — халат (греч. chalaza — бугорок) формируется круговой валик, обрастающий бугорок. Бугорок является нуцеллусом (лат. micella — орешек), а валик развивается в интегумент (лат. integumentum — покров), или покров семязачатка. На вершине семязачатка интегумент не замкнут, поэтому образуется микропиле (греч. mikros — маленький и pyle — вход, отверстие), или пыльцевход. Внутри нуцеллуса выделяется одна клетка и разрастается, становясь клеткой археспория, которая и является материнской клеткой мегаспор. В результате мейоза из нее образуются четыре гаплоидные мегаспоры, расположенные одна над другой. Следовательно, нуцеллус — это мегаспорангий, а семязачаток — видоизмененный спорангий. Мегаспоры из спорангия не высыпаются, одна из четырех прорастает внутри нуцеллуса и формирует женский заросток, остальные отмирают и используются на питание оставшейся. Женский гаметофит внутри семязачатка питается за счет спорофита и представлен многоклеточным телом со значительным количеством запасных веществ, преимущественно масел. На прилегающем к микропиле конце гаметофита образуются два архегония, в брюшке каждого располагается по одной крупной яйцеклетке. Над яйцеклеткой лежит брюшная канальцевая клетка, а в короткой шейке — несколько шейковых канальцевых клеток. На этой стадии гаметофит готов к оплодотворению.
Мужские шишки, или микростробилы, состоят из короткой оси с сидячими на ней микроспорофиллами. Последние представляют собой тонкие пленчатые чешуи с двумя крупными микроспорангиями — пыльцевыми мешками на нижней стороне. Из археспория, расположенного внутри микроспорангия, в дальнейшем образуется тапетум и спорогенная ткань. В результате мейоза из клеток спорогенной ткани формируются многочисленные тетрады гаплоидных микроспор, одевающихся наружной оболочкой — экзиной. У сосны и некоторых других хвойных экзина отстает от внутренней оболочки — интины и образует два воздушных мешка, которые значительно увеличивают парусность микроспоры.
Микроспоры прорастают уже в микроспорангии, формируя мужские заростки, сильно редуцированные и остающиеся в пределах оболочки микроспоры. В результате деления ядра микроспоры вначале отчленяются две маленькие вегетативные протамиальные клетки (лат. protallium — заросток), которые быстро разрушаются. Крупная клетка дает начало двум клеткам: антеридиальной (функционально являющейся антеридием) и сифоногенной, или клетке пыльцевой трубки, часто называемой вегетативной.
Таким образом, мужской заросток первоначально состоит из четырех клеток, но в конце развития включает всего две. На этой стадии микроспорангий вскрывается и микроспоры (пылинки) попадают в воздух. Количество микроспор столь велико, что во время «пыления» сосны они серно-желтой пленкой покрывают листья растений и поверхность водоемов. Переносимые ветром мужские гаметофиты (пылинки) попадают к семязачаткам; так как семенные чешуи женских шишек в это время раздвинуты, происходит опыление. В результате подсыхания микроспора оказывается на поверхности нуцеллуса, сифоногенная клетка формирует пыльцевую трубку. Но в июне в семязачатках еще не сформировался женский заросток с архегониями, а в мужском заростке не образовались мужские гаметы, т. е. сразу после опыления у сосны оплодотворение невозможно. Поэтому заростки продолжают развиваться внутри шишки, чешуи которой сомкнулись. Только после перезимовки произойдет оплодотворение. Но у елей, лиственниц, пихт оплодотворение происходит после опыления. Ему предшествует деление антеридиальной клетки с образованием стерильной клетки (она быстро разрушается) и собственно спермагенной клетки, которая перемещается по пыльцевой трубке, делится кариокинетически и дает безжгутиковые мужские гаметы — спермин. Пыльцевая трубка проникает через нуцеллус в архегоний, спермин выходят из нее и происходит оплодотворение одной яйцеклетки одним из спермиев. Вторая пара гамет не сливается. Оплодотворение происходит внутри семязачатка, и хотя перемещение мужских гаметофитов не нуждается в воде, гаметы находятся в пыльцевой трубке и переносятся ее содержимым (т. е. необходимость сохранения среды с высоким содержанием воды осталась), тем не менее отрыв от водной среды произошел. Такой тип оплодотворения называется сифоногамией (греч. siphon — трубка и gameo — вступаю в брак).
После оплодотворения из семязачатка формируется семя, при этом из зиготы развивается зародыш нового спорофита, ткань заростка разрастается и, обогащаясь питательными веществами, превращается в эндосперм семени (он гаплоиден!). Во времени происходит смена его функций: вначале половая (образование женских гамет), затем запасающая, обеспечивающая питание зародыша за счет спорофита. В этом состоит существенное отличие женского гаметофита голосеменных от заростка папоротников, который живет самостоятельно, автотрофен по типу питания и на первом этапе обеспечивает питание зиготы и зародыша спорофита.
Развитие зародыша и эндосперма ведет к разрушению нуцеллуса, покровы семяпочки превращаются в семенную кожуру; поверхностные ткани семенной чешуи, прилегающие к семени, формируют прозрачное крылышко (у кедровых сосен оно не образуется), облегчающее распространение семян при помощи ветра.
Семя голосеменных — сложное структурное образование: кожура диплоидна и своим происхождением обязана материнскому спорофиту (мегаспорангию); эндосперм гаплоиден и по природе — это женский заросток; диплоидный зародыш — новый спорофит, образующийся в результате оплодотворения из зиготы. Полностью сформированный зародыш имеет хорошо выраженные зачаточные органы: гипокотиль и зародышевый корешок, несколько семядолей (от 5 до 8), прикрывающих зародышевую почечку.
Созревшие семена находятся внутри шишки, защищенные плотно сомкнутыми семенными чешуями, поэтому термин «голосемянность» не совсем точно отражает сущность. Более справедлив термин «голосемяпочковость», так как семяпочки открыто соприкасаются с внешней средой, поэтому пыльца попадает непосредственно на семязачаток (сравните у покрытосеменных!).
Анализируя цикл развития сосны обыкновенной, можно заметить, хотя и тщательно замаскированную, смену поколений с преобладанием спорофита. Раздельнополые гаметофиты гаплоидны, сильно редуцированы и живут за счет спорофита. Краткий этап самостоятельного существования (от высыпания до попадания на семязачаток) имеет только пыльцевое зерно (мужской гамето-фит). Семена же являются одновременно органами расселения (удаляясь на большое расстояние от материнского организма), размножения (на одном растении их образуется огромное количество) и воспроизведения (из каждого семени вырастает новый спорофит, подобный материнскому, но не с идентичной генетической программой).
Поскольку зародыш и семя — результат полового процесса, то справедливо считать семенное размножение одним из типов полового размножения. Биологическая роль семян велика: зародыш надежно защищен семенной кожурой, обеспечен питанием за счет материнского спорофита. Однако являясь более крупным, чем спора, семя требует специальных приспособлений для расселения, что успешно было решено в процессе адаптивной эволюции. Споры же, хорошо распространяемые ветром из-за их незначительных размеров и небольшой массы, более уязвимы и не так надежно обеспечивают процесс размножения. Поэтому образование семян — огромный прогрессивный шаг в эволюционном развитии растений и одно из условий, которое обеспечило семенным растениям широкое распространение на планете в самых различных условиях обитания.
Цветок — это укороченный спороносный побег, в котором «женские» мегаспорофиллы стали плодолистиками, а «мужские» — тычинками. Цве ток имеют только покрытосеменные растения (цветковые), в этом органе протекает спорогенез, гаметогенез, половой процесс.
Источник