2. Филогенетические связи в природе. Естественная классификация живых форм. Основные типы животного мира. Доказательства монофилии.
Филогенетическое дерево (эволюционное дерево, дерево жизни) — дерево, отражающее эволюционные взаимосвязи между различными видами или другими сущностями, имеющими общего предка.
Вершины филогенетического дерева делятся на три класса: листья, узлы и (максимум один) корень. Листья — это конечные вершины, то есть те, в которые входят ровно по одному ребру; каждый лист отображает некоторый вид живых организмов (или иной объект, подверженный эволюции, например, домен белка). Каждый узел представляет эволюционное событие: разделение предкового вида на два или более, которые в дальнейшем эволюционировали независимо. Корень представляет общего предка всех рассматриваемых объектов. Ребра филогенетического дерева принято называть «ветвями».
С позиций общей теории систем наиболее принципиальным представляется деление живых систем по характеру их связей с внешней средой, в соответствии с чем выделяются: 1) открытые живые системы, имеющие обмен веществом с внешней средой, и 2) закрытые живые системы, не имеющие такого обмена.
По уровню функциональной организации живые системы естественно подразделяются на 1) организменные живые системы, как элементарные и неделимые, т.е. отдельные организмы, и 2) надорганизменные живые системы, т.е. различные функционально-структурные, ассоциации организмов — колонии, популяции, виды, экосистемы и т. п.
По уровню структурной агрегации живые системы делятся на три категории: первичные доклеточные организмы — протобионты, б) простые одноклеточные — прокариоты и одноядерные эукариоты и в) вирусы;
метабионтные живые системы, представленные возникшими из мо-нобионтов организмами-метабионтами, у которых генетическая система не централизована в масштабах организма. Таковы многоядерные одноклеточные, ценоцитные и многоклеточные организмы, исключая из числа последних ценометабионтные системы;
ценометабионтные живые системы, представленные возникшими из метабионтов организмами-ценометабионтами, организация которых явилась результатом агрегации метабионтных организменных структур. Таковы системы, традиционно называемые «колониями»: сифонофоры, коралловые полипы, мшанки и т. п.
По способу организации процессов метаболизма живые системы делятся на две группы:
1) автобионтные живые системы, представленные организмами, которые метаболируют самостоятельно, располагая необходимыми для этого собственными энергозапасающими, ферментными и синтезирующими аппаратами; к их числу относятся все клеточные организмы;
2) анавтобионтные живые системы, представленные организмами, которые метаболируют только на основе обязательного использования энергозапасающих, ферментных и синтезирующих аппаратов другого организма
Монофилия (греч. монос — один и филон — род, племя) — это происхождение всего органического мира из единого корня. «Теорию общего происхождения» предложил Ч. Дарвин. В своем произведении он писал: «На основании принципа естественного отбора, сопровождаемого расхождением признаков, не представляется невероятным, что от какой-нибудь низко организованной и промежуточной формы могли развиться как животные, так и растения; а если мы допустим это, мы должны допустить, что и все органические существа, когда-либо жившие на Земле, могут происходить от одной первобытной формы».
3.Общая характеристика п/т хелицероносных. Паукообразные.
Общая характеристика. Наземные членистоногие, ведут родословную от палеозойских водных предков, близких к предкам ракообразных. В расчленении тела нет единообразия. Частый случай, когда сегменты головы и груди, сливаясь, образуют головогрудь. В наибольшей мере слияние сегментов тела представлено у клещей. Антенны отсутствуют, глаза только простые в количестве от одной до восьми пар. Членистых придатков шесть пар. Из них первые две пары: хелицеры и педипальпы — выполняют функцию антенн и челюстей. Остальные четыре пары — ноги. Брюшко обычно лишено ног, но у пауков они видоизменены в паутинные бородавки. У скорпионов ноги на брюшке закладываются в эмбриональном периоде, у взрослых они отсутствуют. В подтипе два класса:Меристомовые — Мечехвосты (Xiphosu-га) и Паукообразные
Паукообразные. в теле паукообразных выделяют два отдела — просому (головогрудь) и опистосому(брюшко). Просома состоит из 6 сегментов, несущих по паре конечностей: хелицеры, педипальпы и четыре пары ходных ног. У паукообразных они несут относительно тонкую хитиновую кутикулу, под которой находится гиподерма ибазальная мембрана. Органами дыхания служат трахеи
Источник
Филогенетическое дерево
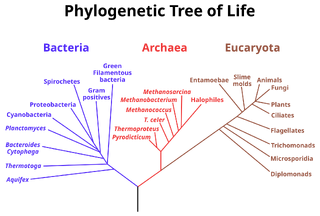
Укоренённое дерево — дерево, содержащее выделенную вершину — корень. Корневое дерево можно считать ориентированным графом, поскольку на нем имеется естественная ориентация — от корня к листьям. Каждый узел корневого дерева отвечает последнему общему предку нижележащих листьев дерева. Рисунок 1 представляет корневое филогенетическое дерево, окрашенное в соответствии с трёхдоменной системой живых организмов. [2] .
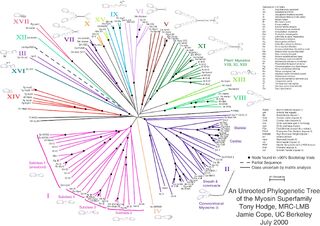
Неукоренённое дерево не содержит корня и отражает связь листьев без предполагаемого положения общего предка. Необходимость рассматривать некорневые деревья возникает из-за того, что часто связи между узлами восстановить легче, чем направление эволюции. Рисунок 2 иллюстрирует некорневое филогетическое дерево. [3] . Наиболее достоверным методом для превращения неукорененного дерева в укорененное (для этого надо либо объявить корнем один из узлов, либо разбить одну из ветвей на две, выходящие из корня) является использование достоверной «внешней группы» видов — достаточно близких к интересующему нас набору видов для достоверного восстановления топологии дерева для объединенного множества видов, но в то же время заведомо являющихся отдельной группой. Иногда положение корня можно угадать, исходя из каких-либо дополнительных знаний о природе изучаемых объектов (видов, белков, etc.)
Укоренённое и неукоренённое филогетическое дерево может быть бифуркационным или небифуркационным, а также маркированным или немаркированным. В бифуркационном дереве к каждому узлу подходят ровно три ветви (в случае корневого дерева — одна входящая ветвь и две исходящие). Таким образом бифуркационное дерево предполагает, что все эволюционные события состояли в происхождении от предкового объекта ровно двух потомков. К узлу небифуркационного дерева могут подходить четыре и более ветви. Маркированное дерево содержит названия листьев, тогда как немаркированное просто отражает топологию.
Дендрограмма — общий термин, обозначающий схематическое представление филогенетического дерева.
Построение филогенетических деревьев [ ]
Филогенетические деревья из неограниченного числа входных последовательностей составляются используя вычислительные филогенетические методы. Методы матричного расстояния, такие как методы ближайшего вхождения, которые требуют множественные выравнивания цепочек для вычисления генетического расстояния, просты в применении; методы выравнивания множества цепочек, наподобие используемых в программе ClustalW выполняют как выравнивание цепочки, так и филогенетических деревьев. Другие методы максимально экономичны и используют приближённые технические приёмы такие, как максимальная вероятность; приближение Байеса также применимо к филогенетике, но оно спорно. [4] Нахождение оптимального дерева, используя многие из этих технических способов НП-полно [5] или НП-трудно [4] , поэтому эвристический поиск и методы [6]
- эффективность (насколько долго вычисление ответа, сколько памяти для этого потребуется?)
- производительность (есть ли польза от полученных данных или информация бесполезна?)
- постоянство (будут ли повторные ответы такими же, если каждый раз даются разные данные для той же проблемной модели?)
- устойчивость к ошибкам (справляется ли с нарушениями в предпосылках расссматриваемой модели?)
- выдача предупреждений (будет ли предупреждать нас, когда неправильно используется, т.е. предпосылки неверные?)
Также методы построения дерева могут быть предложены вниманию математиков. Деревья могут быть построены, используя Т-теорию. [7]
Ограниченность филогенетических деревьев [ ]
См. также [ ]
- ↑ Hodge, T. & M. J. T. V. Cope. 2000. A Myosin Family Tree. Journal of Cell Science113: 3353-3354. See also the Myosin external link below.
- ↑ en:Proceedings of the National Academy of Sciences95: 6854-6859.
- ↑ Maher, B. A. 2002. Uprooting the Tree of Life. The Scientist16: 18 (Sep. 16, 2002); subscription only
- ↑ 4,04,1 Felsenstein J. (2004). Inferring Phylogenies Sinauer Associates: Sunderland, MA.
- ↑ Так говорят о классе комбинаторных задач с нелинейной полиномиальной оценкой числа итераций
- ↑ Penny, D., Hendy, M. D. & M. A. Steel. 1992. Progress with methods for constructing evolutionary trees. Trends in Ecology and Evolution7: 73-79.
- ↑ A. Dress, K. T. Huber, and V. Moulton. 2001. Metric Spaces in Pure and Applied Mathematics. Documenta MathematicaLSU 2001: 121—139
- ↑ Woese, C. R. 2002. On the evolution of cells. Proceedings of the National Academy of Sciences99: 8742-8747.
Изображения в интернете [ ]
- Phylogenetic Trees Based on 16s rDNA
- A 3D View
- Human Y-Chromosome 2002 Phylogenetic Tree
- In 2003, the Science journal dedicated a special issue to the tree of life, including an online version of a tree of life.
Общее описание [ ]
- PhyloCode
- A Multiple Alignment of 139 Myosin Sequences and a Phylogenetic Tree
- Tree of Life Web Project
- http://www.aisee.com/graph_of_the_month/jura.htm The most detailed and comprehensive family tree of dinosaurs yet available
- http://www.omne-vivum.com tree of life with lots of pictures
Эта страница использует содержимое раздела Википедии на русском языке. Оригинальная статья находится по адресу: Филогенетическое дерево. Список первоначальных авторов статьи можно посмотреть в истории правок. Эта статья так же, как и статья, размещённая в Википедии, доступна на условиях CC-BY-SA .
Источник