20. Половое размножение растений
Растения размножаются как бесполым, так и половым способом.
Вопрос 2. Рассмотрите рисунок на с. 139. Что общего и в чём разница в половом размножении хламидомонады и спирогиры?
Общим в половом размножении хламидомонады и спирогиры можно считать образование зиготы, имеющей двойной набор хромосом, которая покрывается плотной оболочкой и в таком состоянии переживает зиму. С наступлением тепла её содержимое делится.
1. Процессы предшествующие образованию зиготы:
При половом размножении хламидомонады под оболочкой материнской клетки развивается много мелких двухжгутиковых гамет (от 32 до 64 клеток). Прорвав оболочку, они выходят наружу, где попарно сливаются с гаметами других особей, образуя зиготы.
Осенью две параллельно расположенные нити спирогиры обволакиваются слизью. В клетках, находящихся одна напротив другой, образуются выросты — мостики, оболочки на концах выростов растворяются. Содержимое одной клетки перетекает в другую, их ядра сливаются — происходит оплодотворение.
2. Процессы, происходящие после образования зиготы:
С наступлением тепла содержимое зиготы хламидомонады делится путём мейоза, образуя четыре хламидомонады. Они выходят наружу и начинают вести самостоятельный образ жизни.
Переждав неблагоприятные условия, ядро зиготы спирогиры дважды делится, три ядра из четырёх отмирают, а одноядерная клетка прорастает в новую нить спирогиры.
Вопрос 3. Почему для полового размножения мхов и папоротников нужна вода, а для цветковых растений — нет?
Мхи и папоротники не образуют пыльцу, надежно защищенную от высыхания. Их мужские половые клетки, спермии, добираются до яйцеклеток вплавь, а на воздухе неподвижны и беззащитны.
Много воды для оплодотворения не требуется. Мхам, например, достаточно тонкой пленки, образованной каплями росы или дождя, но в сухую погоду у них могут возникнуть проблемы с размножением, особенно если мужские и женские растения удалены друг от друга. В этих условиях перенос спермиев могут обеспечить только животные.
Вопрос 4. Чем оплодотворение у цветковых растений отличается от подобного процесса у других организмов?
У цветковых растений в оплодотворении участвуют два спермия, поэтому оплодотворение у них называют двойным. Один из спермиев сливается с яйцеклеткой, образуя зиготу, а второй — с ядром центральной клетки, составляющей большую часть зародышевого мешка.
Вопрос 5. Составьте схему двойного оплодотворения, которая отражала бы суть процесса. Почему оплодотворение называют двойным?
У цветковых растений в оплодотворении участвуют два спермия, поэтому оплодотворение у них называют двойным.
Схема двойного оплодотворения
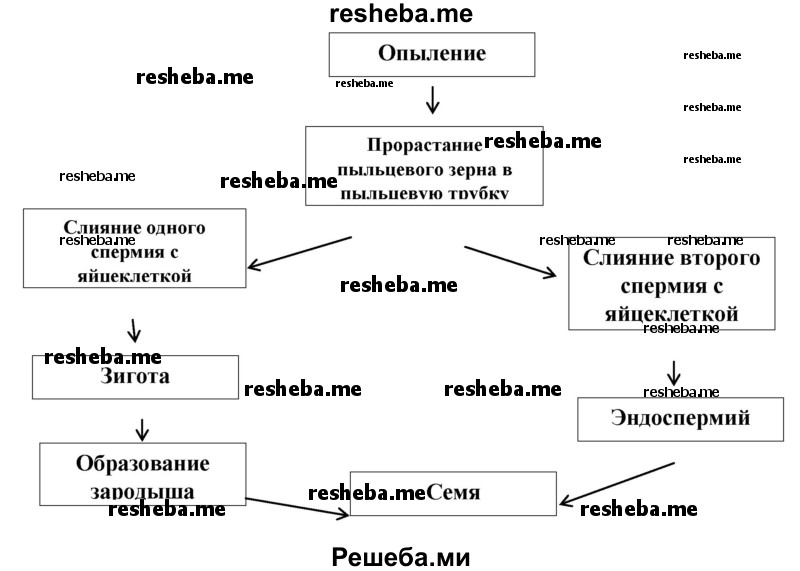
Вопрос 6. Каково биологическое значение двойного оплодотворения?
Несомненное его преимущество очень быстрое (опережающее развитие зародыша) образование питательной ткани, которое происходит только после оплодотворения. Двойное оплодотворение ускоряет весь процесс формирования семяпочки и семени.
Вопрос 7. Какова роль опыления в размножении?
Опыление — перенос пыльцы с тычинок на рыльце пестика. В противном случае гаметы не встретятся и оплодотворения не произойдет.
Вопрос 8. Сравните понятия «перекрёстное опыление» и «самоопыление».
При самоопылении пыльца попадает с тычинки на пестик того же самого цветка. Пшеница, рис, овёс, горох — самоопыляющиеся растения.
При перекрёстном опылении пыльца с тычинок цветка переносится на рыльце пестика цветка другого растения. Различают насекомоопыляемые и ветроопыляемые растения. Насекомоопыляемые растения имеют красивые, яркие цветки или мелкие цветки, собранные в соцветия. Обычно они богаты нектаром, пыльцой, обладают приятным запахом. У ветроопыляемых растений мелкие, невзрачные цветки, собранные в соцветия. Пыльца у них сухая, мелкая, лёгкая. Ветром опыляются тополь, ольха, дуб, рожь.
Источник
5. Двойное оплодотворение цветковых растений
Суть двойного оплодотворения у цветковых растений заключается в том, что в нём участвуют два спермия. Один из них оплодотворяет яйцеклетку, и образуется зигота. Второй спермий сливается с центральной клеткой, из которой развивается запасающая ткань (эндосперм).

Мужской гаметофит (пыльцевое зерно) образуется в пыльцевых камерах пыльников тычинки из микроспоры. Пыльцевое зерно состоит из двух гаплоидных клеток: вегетативной и генеративной, покрытых оболочкой.
Образование женского гаметофита (зародышевого мешка) происходит в завязи пестика в семязачатке из мегаспоры. В состав зародышевого мешка входит семь клеток: гаплоидная яйцеклетка, центральная диплоидная клетка и пять вспомогательных гаплоидных клеток.
При попадании пыльцевого зерна на рыльце пестика начинается деление вегетативной клетки и образуется пыльцевая трубка. Пыльцевая трубка прорастает через столбик пестика и проникает в семязачаток через пыльцевход.
Генеративная клетка пыльцевого зерна делится и образует два спермия. По пыльцевой трубке спермии проникают в семязачаток. Один спермий сливается с яйцеклеткой и образует диплоидную зиготу. Второй спермий сливается с центральной клеткой и образует триплоидную клетку.
Зигота делится, и развивается в зародыш нового растения. Из триплоидной клетки формируется эндосперм. Стенки семязачатка становятся семенной кожурой. Таким образом, семязачаток становится семенем.
Источник
6. Процесс двойного оплодотворения покрытосеменных трастений,его биологическое и эволюционное значение
В завязь покрытосеменных растений проникает два спермия, один из них сливается с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с центральной диплоидной клеткой. Образуется триплоидная клетка, из которой возникнет эндосперм — питательный материал для развивающегося зародыша . Значение двойного оплодотворения,заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения. Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
У цветковых растений имеется ряд особенностей образования половых клеток и оплодотворения. Оплодотворению у них предшествует образование сильно редуцированного гаплоидного поколения — гаметофитов. После оплодотворения прорастание пыльцы цветковых растений начинается с разбухания зерна и образования пыльцевой трубки , которая прорывает спородерму в более тонком ее месте — так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика , в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия . В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий ( нуцеллус ) через микропиле семязачатка , реже — иным образом. Проникнув в зародышевый мешок , пыльцевая трубка разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, и образуется диплоидная зигота, дающая затем начало зародышу . Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка , что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Весь этот процесс получил название двойного оплодотворения. Прочие клетки зародышевого мешка — антиподы и синергиды в оплодотворении не участвуют и довольно быстро разрушаются.
Как и у других высших растений , при мейозе у цветковых образуются споры . Микроспоры (мужские споры) образуются в пыльниках , из них формируются пыльцевые зерна (мужские гаметофиты). Зрелое пыльцевое зерно состоит из трех клеток — вегетативной (или клетки, пыльцевой трубки ) и двух спермиев (лишенные жгутика сперматозоиды). Мегаспоры (женские споры) формируются в семяпочках внутри завязи пестика .
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных , где довольно мощный гаплоидный эндосперм развивается независимо от процесса оплодотворения, у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение же уровня плоидности эндосперма до 3n, по-видимому, способствует более быстрому росту этой полиплоидной ткани по сравнению с диплоидными тканями спорофита .
Взаимодействие пыльцевой трубки гаметофита с тканями спорофита — сложный процесс, регулируемый химическими веществами. Так, выяснилось, что если промыть пыльцу дистиллированной водой, она теряет способность к прорастанию. Если сконцентрировать полученный раствор и обработать концентратом пыльцу, она вновь станет полноценной. После прорастания рост пыльцевой трубки контролируется тканями пестика. Например, у хлопчатника рост трубки до яйцеклетки занимает 12-18 ч, но уже через 6 ч можно установить, к какой семяпочке направляется пыльцевая трубка: в этой семяпочке начинается разрушение особой клетки – синергиды. Во многих случаях у цветковых растений существует «запрет» на самоопыление: спорофит «узнает» своего мужского гаметофита и не разрешает ему участвовать в оплодотворении. В некоторых случаях при этом собственная пыльца не прорастает на рыльце пестика. В большинстве же случаев рост пыльцевой трубки начинается, но затем останавливается и она не достигает яйцеклетки.
Опыление эффективно только при переносе пыльцы с одной формы на другую. За распознавание своей пыльцы отвечают молекулы-рецепторы, представляющие собой сложные комплексы белков с углеводами. Показано, что растения дикой капусты , которые не вырабатывают в тканях рыльца молекул рецептора, могут самоопыляться. У нормальных растений рецепторы появляются на рыльце за день до открытия цветка. Если раскрыть бутон и нанести на него собственную пыльцу за два дня до распускания, то оплодотворение произойдет, а если за один день до распускания — то нет.
В некоторых случаях самонесовместимость пыльцы у растений определяется серией множественных аллелей одного гена, сходно с несовместимостью при пересадках тканей у животных. Эти аллели обозначаются буквой S, и число их в популяции может достигать десятков и даже сотен. Если, например, генотип производящего яйцеклетки растения — s1s2, а производящего пыльцу — s2s3, то прорастать при перекрестном опылении будут только 50% пылинок — те, что несут аллель s3. При наличии десятков аллелей большая часть пыльцы при перекрестном опылении нормально прорастает, а самоопыление полностью предотвращается.
Втори́чный эндоспе́рм — ткань, образующаяся в семенах большинства цветковых растений во время оплодотворения. Эндосперм окружает зародыш и обеспечивает его питание за счет крахмала, растительных масел и белков. Это делает эндосперм цветковых растений важным источником питательных веществ в диете человека. Например, эндосперм пшеницы после размалывания представляет собой муку, из которой пекут хлебобулочные изделия, эндосперм ячменя используют для пивоварения.
Первичный эндосперм голосеменных образуется до оплодотворения из мегаспоры и соответствует женскому гаметофиту. Клетки эндосперма голосеменных первоначально гаплоидные, затем в результате слияния ядер становятся полиплоидными.
Происхождение эндосперма у покрытосеменных:
Примитивные цветковые растения имеют семена с крупным эндоспермом и мелким зародышем. Эволюционное развитие привело к появлению растений с семенами, в которых эндосперм невелик или отсутствует. У продвинутых групп цветковых зародыш занимает большую часть семени, и эндосперм не развивается или потребляется зародышем к моменту созревания семени.
Древнее цветковое растение Nuphar polysepala имеет диплоидный эндосперм, результат слияния спермия с одной полярной клеткой. Считается, что на ранней стадии развития цветковых произошло изменение в данном типе развития и стал образовываться не семи-, а восьмиклеточный зародышевый мешок с диплоидным эндоспермом.
Существует два типа образования эндосперма — ядерный (нуклеарный, nuclear), при этом образуется жидкий эндосперм и клеточный, когда происходит образование клеточных стенок при делении ядер. Среди покрытосеменных наиболее распространен клеточный тип. Сладкая кукуруза собирается в период ядерного образования эндосперма, до момента превращения сахаров в крахмал и формирования клеточных стенок. Кокосовое молоко является жидким эндоспермом.
Значение в развитии растения
Эндосперм осуществляет транспорт питательных веществ из материнского организма в семя, в эндосперме может происходить импринтинг генов, и именно в эндосперме останавливается развитие семян генетически модифицированных растений. У покрытосеменных эндосперм содержит гормоны, например, цитокинины, которые регулируют дифференцировку клеток и образование зародышевых органов.
Источник