Корневая система как орган поглощения воды
Наземные растения, в основном, поглощают воду из почвы. Однако некоторое количество воды может попадать в листья из воздуха. Есть даже растения, для которых атмосфера является главным источником влаги. Это эпифиты, живущие на поверхности других растений, но не являющиеся паразитами. Они обладают воздушными корнями с полыми тонкостенными клетками и впитывают парообразную влагу и воду осадков подобно губке. У некоторых эпифитов дождевая вода собирается листьями и затем всасывается с помощью листовых волосков.
Корневая система является органом поглощения воды из почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной структурой. Подсчитано, что общая поверхность корневой системы может превышать поверхность надземных органов примерно в 150 раз. Рост корня и его ветвление продолжаются в течение всей жизни растения.
Поглощение воды и питательных веществ осуществляется корневыми волосками ризодермы. Ризодерма — это однослойная ткань, покрывающая корень снаружи. У одних видов растений каждая клетка ризодермы формирует корневой волосок, у других она состоит из двух типов клеток: трихобластов, образующих корневые волоски, а атрихобластов, не способных к образованию волосков.
Из ризодермы вода попадает в клетки коры. У травянистых растений кора корня обычно представляет собой несколько слоев живых паренхимных клеток. Между клетками имеются крупные межклетники, обеспечивающие аэрацию корня. Через клетки коры возможны два пути транспорта воды и растворов минеральных солей: по симпласту и апопласту. Более быстрый транспорт воды происходит по апопласту, так как в цитоплазме вода отвлекается на нужды клетки.
Затем вода попадает в клетки эндодермы. Эндодерма — это внутренний слой клеток коры, граничащий с центральным цилиндром. Их клеточные стенки водонепроницаемы из-за отложения суберина и лигнина (пояски Каспари). Поэтому вода и соли проходят через клетки эндодермы по симпласту и транспорт воды в эндодерме замедляется (рис. 5). Это необходимо, так как диаметр стели (центрального цилиндра), куда попадает вода из эндодермы, меньше всасывающей поверхности корня.
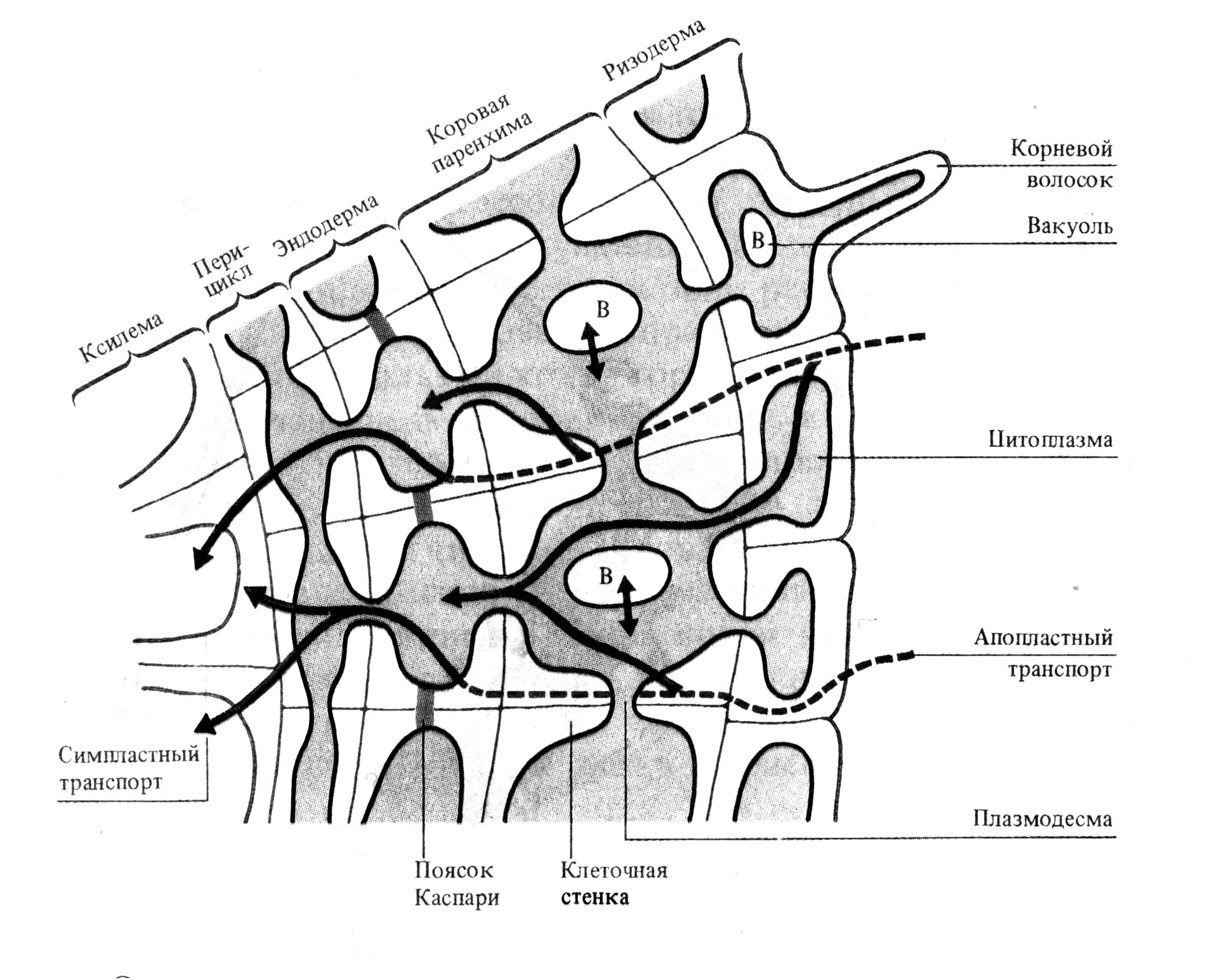
Рис. 5. Схема путей радиального транспорта воды и ионов через корень до сосудов ксилемы (по J. Moorby, 1981 – цит. по В. В. Полевому).
Центральный цилиндр корня содержит перицикл и две системы проводящих элементов: ксилему и флоэму. Клетки перицикла представляют собой одно- или многослойную обкладку проводящих сосудов. Его клетки регулируют транспорт веществ как из наружных слоев в ксилему, так и из флоэмы в кору. Кроме того, клетки перицикла выполняют функцию образовательной ткани, способной продуцировать боковые корни. Паренхимные клетки перицикла активно транспортируют ионы в проводящие элементы ксилемы. Контакт осуществляется через поры во вторичных клеточных стенках сосудов и клеток. Между ними нет плазмодесм. Затем вода и растворенные вещества диффундируют в полость сосуда через первичную клеточную стенку. Для некоторых паренхимных клеток сосудистого пучка характерны выросты — лабиринты стенок, выстланные плазмалеммой, что значительно увеличивает ее площадь. Эти клетки активно участвуют в транспорте веществ в сосуды и обратно и называются передаточными или переходными. Они могут граничить одновре-менно с сосудами ксилемы и ситовидными трубками флоэмы. По сосудам флоэмы транспортируются органические вещества из надземной части растения в корни.
Вода пассивно диффундирует в сосуды ксилемы благодаря осмотическому механизму. Осмотически активными веществами в сосудах являются минеральные ионы и метаболиты, выделяемые насосами плазмалеммы паренхимных клеток, окружающих сосуды. Сосущая сила сосудов выше, чем у окружающих клеток из-за повышающейся концентрации ксилемного сока и отсутствия значительного противодавления со стороны малоэластичных клеточных стенок. В результате поступления воды в сосудах ксилемы развивается гидростатическое давление, получившее название корневого давления. Оно участвует в поднятии ксилемного раствора по сосудам ксилемы из корня в надземную часть растения. Поднятие воды по растению вследствие развивающегося корневого давления называют нижним концевым двигателем.
Примером работы нижнего концевого двигателя служат плач растений. Весной у кустарников и деревьев с еще нераспустившимися листьями можно наблюдать интенсивный ксилемный ток снизу вверх через надрезы ствола и веток. У травянистых растений при отрезании стебля из пенька выделяется ксилемный сок, называемый пасокой.
Поступление воды через корневую систему сокращается с понижением температуры. Это происходит по следующим причинам: 1) повышается вяз-кость воды и поэтому снижается ее подвижность, 2) уменьшается проницаемость протоплазмы для воды, 3) тормозится рост корней, 4) уменьшается скорость метаболических процессов. Поступление воды снижается при ухудшении аэрации почвы. Это можно наблюдать, когда после сильного дождя почва залита водой, но при ярком солнце из-за сильного испарения растения завядают. Большое значение имеет концентрация почвенного раствора. Вода поступает в корень только тогда, когда водный потенциал корня меньше водного потенцииала почвы. Если почвенный раствор имеет более отрицательный потенциал, вода не будет поступать в корень, а выходить из него.
Источник
Вопрос 4. Корень как орган поглощения и усвоения питательных веществ.
Ко́рень (лат. radix) — осевой, обычно подземный вегетативный орган высших сосудистых растений, обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными минеральными веществами к стеблю и листьям.
На корне нет листьев, в клетках корня нет хлоропластов.
Кроме основного корня, многие растения имеют боковые и придаточные корни. Совокупность всех корней растения называют корневой системой. В случае, когда главный корень незначительно выражен, а придаточные корни выражены значительно, корневая система называется мочковатой. Если главный корень выражен значительно, корневая система называется стержневой.
Тело первых вышедших на сушу растений ещё не было расчленено на побеги и корни. Оно состояло из ответвлений, одни из которых поднимались вертикально, а другие прижимались к почве и поглощали воду и питательные вещества. Несмотря на примитивное строение, эти растения были обеспечены водой и питательными веществами, так как имели небольшие размеры и жили около воды.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и дали начало корням, приспособленным к более совершенному почвенному питанию. Это сопровождалось глубокой перестройкой их структуры и появлением специализированных тканей. Образование корней было крупным эволюционным достижением, благодаря которому растения смогли осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх к свету. Например, у мохообразных настоящих корней нет, их вегетативное тело небольших размеров — до 30 см, обитают мхи во влажных местах. У папоротникообразных появляются настоящие корни, это приводит к увеличению размеров вегетативного тела и к расцвету этой группы в каменноугольный период.
Некоторые растения откладывают в корне запасные питательные вещества, такие образования называют корнеплодами.
Корень — один из основных вегетативных органов растения. К его функциям относятся:
поглощение минеральных веществ,
синтез органических веществ,
выделение продуктов обмена,
запасание питательных веществ.
Тонкая оболочка корневого волоска плотно склеивается с комочками почвы. Корневые волоски служат опорой для растущей верхушки корня, выполняют функцию поглощения воды и минеральных веществ из почвы. Длина корневого волоска — 0,6-10 мм, но общая длина волосков весьма значительна и может достигать в сумме 20 км. Волоски выделяют в почву различные вещества, растворяющие труднодоступные растению соединения в почве. Продолжительность жизни корневого волоска — 15-20 дней.
Функционально поглотительной тканью корня являются корневые волоски (у водных растений — ризодермис), а первичная ассимиляция ионов происходит в коре корня.
Корневая система поглощает из почвенного поглощающего комплекса через почвенный раствор все необходимые элементы, причем способна растворять и нерастворимые соединения за счет выделения органических кислот. Это явление подтверждается опытами с использованием мраморной пластинки в сосуде, на которой ясно образуется отпечаток корневой системы растения, растворяющей мрамор в местах соприкосновения с корневыми волосками. Лучшему усвоению минеральных веществ способствует внесение физиологически кислых солей (при этом происходит освобождение анионов из трудно растворимых соединений), например, внесение сульфата аммония в почвы высвобождает фосфорную кислоту из фосфоритов, а внесение натриевой селитры не производит такого действия.
Корневые выделения при бессменном возделывании одной культуры могут влиять на растения отрицательно, создавая неблагоприятную рН почвенного раствора. При этом происходит накопление вредной микрофлоры, возбудителей корневых гнилей, что приводит к увеличению степени поражаемости культуры болезнями.
Корневые выделения состоят из:
веществ, которые отчуждаются клетками в обмен на поглощаемые ионы питательных солей,
веществ, теряемых корнем вследствие выщелачивания наружным раствором и » вытягиваемых» их корня электростатическими силами,
веществ, которые освобождаются отмирающими клетками чехлика, эпидермиса и коры.
Корневая система растений — это и место синтеза многих важных органических соединений, таких как аминокислоты, алкалоиды, гормоны и ряд других веществ. О синтетической роли корневой системы свидетельствует факт прекращения жизнедеятельности срезанных растений, даже если их помещают в питательный раствор. Только если у срезанного побега образуются новые придаточные корни, рост растения возобновляется. В культуре in vitro рост корневой ткани неограничен, а культуру клеток из стебля получить не удается.
Количественные показатели поглощения минеральных элементов.
Между клетками корня могут существовать значительные электрические градиенты, которые возникают и исчезают, по-видимому, вследствие включения и выключения ионных насосов и изменения концентрационных градиентов в тканях корня.
Поглощенные ионы должны сначала пересечь плазмалемму, чтобы проникнуть в цитоплазму, а затем и в мембрану, окружающую вакуоль, или другую органеллу, чтобы попасть затем во внутренний компартмент цитоплазмы.
Ионы могут проникать через мембрану активно или пассивно, при этом они перемещаются благодаря своей кинетической энергии, не потребляя при этом энергию АТФ. Липидорастворимые молекулы проходят сквозь мембраны легко, многие неорганические ионы, не растворимые в липидах, проникают, по-видимому, через специальные водные белковые каналы в мембране, то есть используется механизм облегченной диффузии с участием пермеаз.
Источник