Печёночные мхи
Печёночные мхи — мелкие и нежные мохообразные растения.
Одни из них снабжены стебельками и листьями, лишенными всяких жилок и расположенными в два или три ряда; те, что находятся на стороне, обращенной к почве, чешуевидны и совершенно другой формы, чем остальные. Верхние листья, располагаясь обыкновенно в два ряда, имеют две лопасти, из которых одна маленькая принимает особую форму и пригнута книзу. Таковы листостебельные печёночницы.
Другие представляют плоское или плосковатое слоевище, распростёртое на земле или даже плавающее на воде. Это слоевище обыкновенно ветвится развилисто, тёмно-зелёного цвета и несёт у некоторых, на нижней стороне, нежные чешуйки, расположенные в два ряда и соответствующие, очевидно, листьям.
Мужские (антеридии) и женские (архегонии) органы этих растений построены совершенно наподобие тех же органов лиственных мхов, но расположены они часто иначе, иначе и развиваются после оплодотворения. У листоносных они, подобно лиственным мхам, занимают верхушки стебельков, у слоевцовых — или образуются особые отростки разной формы, несущие на верхушке органы оплодотворения, или же эти органы сидят на самом слоевище. После оплодотворения нижняя вздутая часть архегония, разрастаясь, не выносится, как у лиственных мхов, вверх, а пробуравливается развивающимся спороплодником и остаётся при основании его ножки в виде нежного влагалища. Самый спороплодник раскрывается створочками или зубцами на 2, 4 и больше; у многих он содержит, кроме спор, так называемые элатеры, то есть веретенчатые, извилистые клеточки, содержащие спиральное утолщение внутри; элатеры, вследствие своей гигроскопичности, действуют как пружинки, способствуя раскрыванию спор.
Кроме полового размножения, печеночные мхи способны размножаться с помощью особых почек, содержащихся, как, например, у маршанции, в особых блюдцевидных вместилищах. Заросток у этих растений незначительный, а корни заменены одноклетными волосками, обильно образующимися на нижней стороне слоевища или на основании стебельков.
Юнгерманниевые печёночники [ ]
У юнгерманниевых печёночников (Jungermanniales) стебель построен из одинаковых клеток, например у рода Слоевищные печеночники [ ]
У разных представителей слоевищных печеночников наблюдаются значительные различия в размерах, формах роста и общем облике. Наиболее часто встречаются плоские, стелющиеся по земле, дихотомически ветвящиеся слоевища, но иногда они могут выглядеть как обособленные округлые розетки разной величины. В субтропических и тропических странах наблюдается особенно сложная дифференциация гаметофоров, если в нижней части они приобретают вид корней и стеблей, а в верхней сохраняют листовидную форму с различным характером расчленения пластинки. Это придает таким печеночникам своеобразную уплощенно древовидную форму. Наиболее простой тип слоевища можно видеть у печеночников из рода сферокарпус (Sphaerocarpus. Дальнейшее усложнение строения слоевища наблюдается у паллавицинии (Pallavicinia) и близких к ней родов симфиогины (Symphyogутф), мерчии (Moerckia) и гименофитума (Нуmenophytum), у которых в основной ткани слоевища проходит один или два проводящих пучка вытянутых клеток. В центре каждого пучка находится группа гидроидов-пористых клеток с утолщенными продольными стенками, проводящих воду. Гидроиды окружены двумя-тремя рядами лептоидов — тонкостенных, содержащих плазму, вытянутых в длину и разделенных косыми поперечными стенками клеток. По лептоидам осуществляется передвижение органических веществ. У видов рода метцгерия (Metzgeria) слоевище представлено многослойной жилкой, одетой крупными эпидермальными клетками и отходящей от неё однослойной пластинкой. Усложнение строения слоевища проявляется далее в расчленении пластинки на отдельные листовидные лопасти и в появлении разнообразных чешуек на нижней или верхней стороне слоевища. Сложное строение слоевища характерно для представителей порядка маршанциевых (Marchantiales). У наземных видов рода риччия (Riccia) слоевище состоит из двух тканей: нижней — основной и верхней — ассимиляционной. Основная ткань образована несколькими слоями тонкостенных бесхлорофилльных паренхимных клеток, в которых откладывается крахмал. С нижней стороны основная ткань покрыта одним—тремя слоями эпидермальных клеток, от которых отходят ризоиды и брюшные чешуйки, расположенные в один или два ряда. Ассимиляционная ткань построена из однорядных вертикальных столбиков паренхимных клеток с хлоропластами. Одна или две верхние клетки столбиков (не содержащие хлоропластов) более-крупные. Они закрывают проход в воздушные каналы между столбиками клеток, что защищает слоевище от высыхания. У водных риччий ассимиляционная ткань образует воздушные камеры.
Маршанциевые печеночники [ ]
У маршанциевых печеночников слоевище дифференцировано. Основная ткань состоит из нескольких слоев паренхимных клеток, содержит крахмал, иногда масляные тельца, а также клетки и каналы, заполненные слизью. Снизу она покрыта однослойной эпидермой. Иногда в основной ткани, встречаются буроокрашенные волокна, а у коноцефалума (Conocephalum) — клетки со штриховатыми утолщениями стенок. Ассимиляционная ткань разделена на воздушные камеры, которые отделены одна от другой однослойными стенками и покрытые сверху однослойной эпидермой. Воздушные камеры наверху открываются устьицами. Простые устьица окружены одним или несколькими кругами замыкающих клеток по 4—8 клеток в ряду. бабоочонковидные устьица по форме очень похожи на каналы, образованные несколькими расположенными друг над другом кольцами клеток. У некоторых представителей в воздушных камерах отсутствуют ассимиляционные нити, и их функцию выполняют богатые хлоропластами клетки стенок камер. Разнообразие листостебельных печеночников в основном зависит от чрезвычайного варьирования формы и размеров филлидиев, обычно называемых для удобства листьями. На округлом, большей частью моноподиально разветвленном и обычно лежачем стебле листья расположены в два или три ряда. Сходные своими очертаниями боковые листья располагаются по обе стороны стебля, снизу расположенн один ряд нижних (брюшных) листьев, называемых амфигастриями. Амфигастрии значительно отличаются от боковых листьев формой и величиной.
Литература [ ]
- Абрамов И. И., Абрамова А. Л. Класс печёночники, или печёночные мхи (Marchantiopsida, или Hepaticopsida) // Жизнь растений. В 6-ти т. / Гл. ред. Ссылки [ ]
Проверено 19 декабря 2008 г.
Источник
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса: Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida)
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha) (рис 11. 1).

Рис. 11. 1. Цикл воспроизведения маршанции: 1 – таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6 – вертикальный разрез через женскую подставку (а – архегоний).
Гаметофит имеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами. На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии. На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).
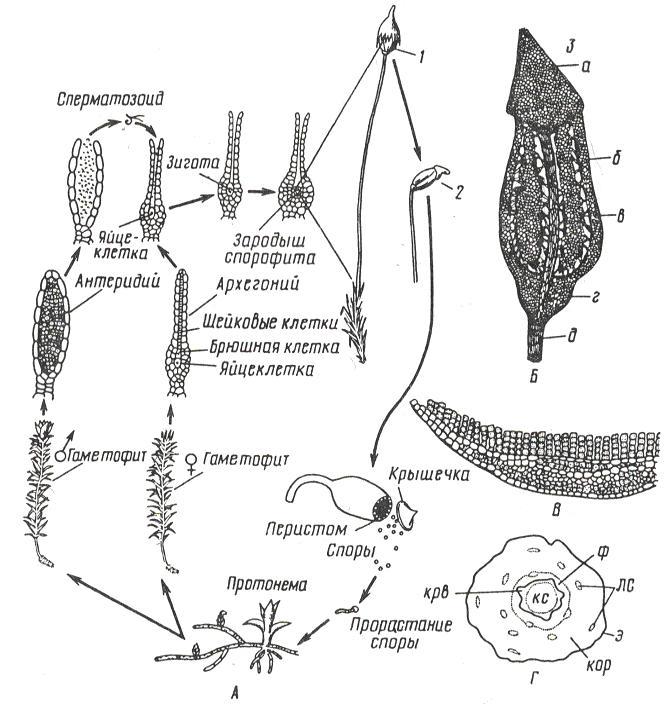
Рис. 11. 2. Кукушкин лен: А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum) (рис. 11. 3).
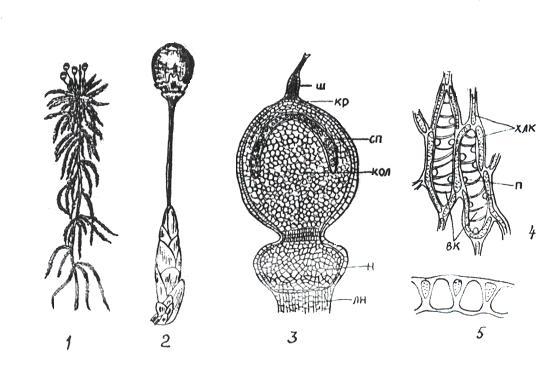
Рис 11. 3. Сфагнум: 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон ( ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа ( хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Источник