Влияние на растения недостатка влаги.
Недостаток воды в тканях растений возникает в результате превышения ее расхода на транспирацию перед поступлением из почвы. Это часто наблюдается в жаркую солнечную погоду к середине дня. При этом содержание воды в листьях снижается на 25—28 % по сравнению с утренним, растения утрачивают тургор и завядают. В результате снижается и водный потенциал листьев, что активизирует поступление воды из почвы в растение.
Различают два типа завядания: временное и глубокое. Причиной временного завядания растений обычно бывает атмосферная засуха, когда при наличии доступной воды в почве она не успевает поступать в растение и компенсировать ее расход. При временном завядании тургор листьев восстанавливается в вечерние и ночные часы. Временное завядание снижает продуктивность растений, так как при потере тургора устьица закрываются
и фотосинтез резко замедляется. Наблюдается, как отмечал А. Г. Лорх, «простой» растений в накоплении урожая.
Глубокое завядание растений наступает, когда в почве практически нет доступной для корней воды. Происходит частичное, а при длительной засухе и общее иссушение и даже гибель растительного организма. Характерный признак устойчивого водного дефицита — сохранение его в тканях утром. Временное и даже глубокое завядание может рассматриваться как один из способов защиты растения от летального обезвоживания, позволяющих некоторое время сохранять воду, необходимую для поддержания жизнеспособности растения. Завядание может происходить при разной потере воды растениями: у тенелюбивых — при 3—5 %, у более стойких — при водном дефиците в 20 и даже 30 %.
Водный дефицит и завядание в разной мере влияют на физиологическую деятельность растения в зависимости от длительности обезвоживания и вида растения. Последствия водного дефицита при засухах многообразны. В клетках снижается содержание свободной воды, возрастает концентрация и снижается рН ваку-олярного сока, что влияет на гидратированность белков цитоплазмы и активность ферментов. Изменяются степень дисперсности и адсорбирующая способность цитоплазмы, ее вязкость. Резко возрастают проницаемость мембран и выход ионов из клеток, в том числе из листьев и корней (экзоосмос); эти клетки теряют способность к поглощению питательных веществ.
При длительном завядании снижается активность ферментов, катализирующих процессы синтеза, и повышается ферментов, катализирующих гидролитические процессы, в частности распад (протеолиз) белков на аминокислоты и далее до аммиака, поли-сахаридов (крахмала на сахара и др.), а также других биополимеров. Многие образующиеся продукты, накапливаясь, отравляют организм растения. Нарушается аппарат белкового синтеза. При возрастании водного дефицита, длительной засухе нарушается нуклеиновый обмен, приостанавливается синтез и усиливается распад ДНК. В листьях снижается синтез и усиливается распад всех видов РНК, полисомы распадаются на рибосомы и субъединицы. Прекращение митоза, усиление распада белков при прогрессирующем обезвоживании приводят к гибели растения.
Безусловно, происходящие изменения до определенного этапа в условиях обезвоживания играют и защитную роль, приводят к увеличению концентрации клеточного сока, снижению осмотического потенциала, а следовательно, повышают водоудерживаю-щую способность растения. При недостатке влаги суммарный фотосинтез снижается, что является следствием в основном недостатка СО2 в листьях; нарушения синтеза и распада хлорофил-лов и других пигментов фотосинтеза; разобщения транспорта электронов и фотофосфорилирования; нарушения нормального хода фотохимических реакций и реакций ферментативного вое-
становления СО2; нарушения структуры хлоропластов; задержки оттока ассимилятов из листьев. По данным В. А. Бриллиант (1925), уменьшение оводненности листа у сахарной свеклы на 3—4 % приводит к снижению фотосинтеза на 76 %.
При нарастающем обезвоживании у незасухоустойчивых растений в первый период завядания интенсивность дыхания возрастает возможно из-за большого количества простых продуктов (гексоз) гидролиза полисахаридов, в основном крахмала, а затем постепенно снижается. Однако выделяющаяся в процессе дыхания энергия не аккумулируется в АТФ, а рассеивается в виде теплоты (холостое дыхание). При действии на растения высокой температуры (45 °С) и суховея происходят глубокие структурные изменения митохондрий, повреждение или ингибирование ферментов фосфорилирующего механизма. Все это свидетельствует о нарушении энергетического обмена растений. В корнях и пасоке повышается содержание амидов. В результате тормозится рост растения, особенно листьев и стеблей, снижается урожай. У более засухоустойчивых растений все эти изменения менее выражены.
Из физиологических процессов наиболее чувствительным к недостатку влаги является процесс роста, темпы которого при нарастающем недостатке влаги снижаются значительно раньше фотосинтеза и дыхания. Ростовые процессы задерживаются даже после восстановления водоснабжения. При прогрессирующем обезвоживании наблюдается определенная последовательность в действии засухи на отдельные части растения.
Если рост побегов и листьев в начале засухи замедляется, то корней даже ускоряется и снижается лишь при длительном недостатке воды в почве. При этом молодые верхние по стеблю листья оттягивают воду от более старых нижних, а также от плодоэлементов и корневой системы. Отмирают корни высоких порядков и корневые волоски, усиливаются процессы опробковения и суберинизации. Все это приводит к сокращению поглощения корнями воды из почвы. После длительного завядания растения оправляются медленно и функции их полностью не восстанавливаются. Затянувшееся завядание при засухе приводит к резкому снижению урожая сельскохозяйственных культур или даже к их гибели. При внезапном и сильном напряжении всех метеорологических факторов растение может быстро погибнуть в результате высыхания (захват) или высоких температур (запал). Засухоустойчивость различных органов растений неодинакова. Так, молодые растущие листья за счет притока ассимилятов дольше сохраняют способность к синтезу, относительно более устойчивы, чем листья, закончившие рост, или старые, которые при засухе подвядают в первую очередь.
В условиях затянувшейся засухи отток воды и веществ в молодые листья может происходить и из генеративных органов.
Засуха в ранние периоды развития приводит к гибели цветочных зачатков, их стерильности (белоколосица), а в более поздние — к образованию щуплого зерна (захват). При этом захват будет более вероятен при хорошо развитой к началу засухи листовой поверхности. Поэтому при сочетании влажной весны и начала лета с очень сухой второй половиной (или даже отдельных сильных суховеев) опасность снижения урожая наиболее вероятна.
Источник
7.1. Водный дефицит и устойчивость к засухе
Недостаток воды в растительных тканях возникает тогда, когда расход начинает превышать поступление. Засухи бывают почвенные и атмосферные. Атмосферная засуха характеризуется низкой относительной влажностью воздуха; почвенная засуха, как правило, следует за атмосферной и возникает при длительном отсутствии дождей. Водный дефицит вызывает серьезные нарушения в первую очередь процессов роста и фотосинтеза. Во время засухи наряду с обезвоживанием происходит перегрев растений, поэтому засуха является наиболее жестким стрессовым воздействием. В зонах, где среднегодовое количество осадков позволяет получать очень высокие урожаи сельскохозяйственных культур, главным фактором, лимитирующим рост растений, является дефицит воды. Более устойчивы к засухе такие виды растений, которые способны расходовать воду более экономно или запасать ее.
У растительных организмов имеется несколько адаптивных стратегий, с помощью которых удается переживать засушливые периоды. Одни виды растений способны накапливать и удерживать воду, поддерживая, таким образом, необходимую степень гидратации своих клеток и органов (например, суккуленты, имеющие САМ-путь углеродного метаболизма). Другие приспособились функционировать в условиях дефицита воды в организме (ксерофиты, для которых, как правило, характерен С4-путь фотосинтеза). У некоторых видов растений в условиях засухи (или засоления) даже индуцируется переход фотосинтеза к САМ-пути углеродного метаболизма. К третьей группе относят растения, которые «избегают» засухи за счет организации своего жизненного цикла таким образом, чтобы вегетировать в период достаточного обеспечения водой.
Многие виды растений резко ускоряют свой рост весной, когда в почве накапливается большое количество влаги, до наступления засушливого лета. У таких растений формируется большая листовая поверхность и быстро развивается мощная корневая система, которая выбирает всю влагу из почвы, накопившуюся в зимний период. При этом растения или быстро завершают свой жизненный цикл (эфемероиды), или запасают воду для завершения репродуктивного этапа развития. Такой тип роста и развития растений называют детерминированным, поскольку он определяется периодами дождей или засухи. При резком наступлении засушливого периода растения, как правило, сбрасывают часть листьев за счет формирования у черешка отделительного слоя под влиянием фитогормона этилена. Если водный дефицит развивается постепенно, то растительному организму лучше удается приспособиться к недостатку воды, заранее замедлив темпы ростовых процессов.
Совсем другая стратегия развития у растений, произрастающих в условиях нормальной обеспеченности водой. В этом случае развиваются большие листья, а вегетация и цветение идут весь летний период. Такой тип развития растений называют недетерминированным. У растений с этим типом роста дефицит воды лимитирует не только размеры отдельных листьев, но и их количество.
Одной из самых ранних реакций растительных организмов на водный дефицит является синтез фитогормона абсцизовой кислоты, которая вызывает закрывание устьиц и таким образом снижает потерю воды при транспирации. При водном дефиците транспорт ассимилятов из листьев угнетается в меньшей степени, чем их синтез. Недостаток воды инициирует рост корней в более влажные участки почвы при условии, что в растущие кончики корней поступает необходимое количество ассимилятов. В этом случае возникает конкуренция за ассимиляты между растущими корнями и формирующимися плодами. В условиях водного дефицита активируется синтез низкомолекулярных осмотически активных веществ (моно- и олигосахаридов, аминокислот, в первую очередь пролина, бетаина, многоатомных спиртов) и различных стрессовых белков, таких, как осмотин, дегидрины, а также белков, удерживающих воду и ионы.
В состав стрессовых белков входят аквапорины – каналоформеры, которые облегчают водный транспорт через мембраны в осмотически зависимых ситуациях. У растений найдены два типа аквапорины – в плазматической и вакуолярной мембранах.
Особое значение в осморегуляции имеет пролин, содержание которого при засухе резко возрастает. К функциональным белкам, способствующим устойчивости к засухе, относятся и многочисленные гидролазы, в том числе ферменты протеолиза, ингибиторы протеаз, ферменты биосинтеза осмотиков. При водном стрессе синтезируются также регуляторные белки, которые участвуют в экспрессии генов и сигнальной трансдукции. Пути приспособления растений к засухе приведены на рис. 7.2.
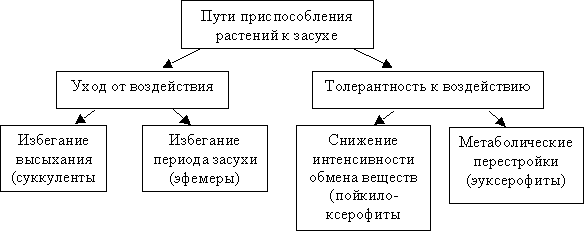
Рис. 7.2. Пути приспособления растений к засухе
Источник