Основные типы устьичного аппарата
Разнообразие типов устьичного аппарата привело к необходимости их классификации. Разные типы устьичного аппарата различаются прежде всего наличием побочных клеток, их числом и расположением по отношению друг к другу и к замыкающим клеткам (рис. 4 и 5).
Наряду с классификациями морфологических или топографических типов устьичного аппарата за последнее десятилетие появились классификации онтогенетических их типов. Первая попытка классифицировать онтогенетическиетипы была предпринята индийским ботаником Д. Пантом в 1965 г. Он выделил три крупные категории устьичных аппаратов в зависимости от того, происходят ли клетки, окружающие устьице, из той же самой материнской клетки, что и устьице, или из других клеток:
1) мезогенный (от греч. mesos — средний и genos — происхождение) устьичный аппарат, у которого все побочные клетки или одна кольцеобразная побочная клетка происходит из той же самой материнской клетки, что и замыкающие клетки;
2) мезоперигенный устьичный аппарат, у которого только одна из окружающих клеток образуется таким путем, остальные образуются из других клеток;
3) перигенный (от греч. peri — около,, кругом) устьичный аппарат, у которого ни одна из окружающих клеток не происходит и» той же самой материнской клетки, что и замыкающие клетки.
Таким образом, в основу классификации Панта был положен способ развития устьичного аппарата без учета типов взрослых структур, тогда как классификация морфологических типов строилась исключительно на расположении побочных клеток в зрелом состоянии, при полном игнорировании способа развития устьичного аппарата. Только появление в 1973 г. классификации онтогенетических типов устьичного аппарата, разработанной Е. Фринс-Классенсом и В. Ван-Коттемом, устранило разрыв между морфологической и онтогенетической классификациями.
С эволюционной точки зрения наиболее примитивным среди высших растений является аномоцитный тип устьичного аппарата: он обнаружен у первых их представителей — риниофитов. Устьица у современных мхов и антоцеротовых также обычно аномоцитные, хотя у некоторых мхов устьица окружены кольцом из более или менее дифференцированных клеток (табл. 1 и 2).
Однако не во всех группах высших растений эволюция шла в направлении от устьичного аппарата, лишенного побочных клеток, к различным типам устьиц с побочными клетками. У цветковых примитивным типом устьичного аппарата является парацитный, тогда как аномоцитный произошел от устьичного аппарата с побочными клетками.
Совсем недавно парацитный устьичный аппарат был обнаружен у одного из палеозойских семенных папоротников — алетоптериса Салливанта (Alethopteris sullivantii). Это представляет значительный интерес, поскольку семенные папоротники принято считать вероятными предками цветковых растений.
В эволюции онтогенетического типа устьичного аппарата голосеменных исходным является перигенный тип, а у цветковых примитивным является мезогенный устьичный аппарат.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Источник
1.8. Типы устьичных клеток
В литературе описывается 19 типов [1], нами выбраны только те, которые используются в анализе лекарственного растительного сырья**.
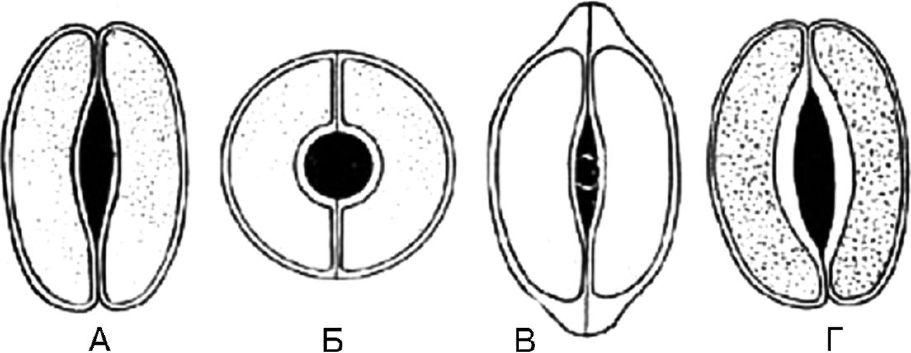
Рис. 63. Типы устьичных клеток. А — чечевицевидные; Б — сферовидные; В — колпачковидные; Г – ладьевидные
1.8.1. Чечевицевидные — 2 одинаковые клетки полулунной формы расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель веретеновидная (рис. 63, А). Тип устьичных клеток характерен для большинства растений.
1.8.2. Сферовидные — две одинаковые, сильно кругообразноизогнутые клетки расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель круглая (рис. 63, Б).
1.8.3.Колпачковидные — две одинаковые клетки полулунной формы в полярных частях имеют утолщения в виде колпачка. Щель веретеновидная (рис. 63, В). Встречаются у наперстянок.
1.8.4.Ладьевидные — внутренние стенки устьичных клеток утолщены. Щель веретеновидная (рис. 63, Г). Наблюдается в траве золототысячника, в листьях вахты.
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток происходит активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза, сахарами влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток. В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие, что ведет к увеличению объема последних и резкому возрастанию тургора. Утолщение «брюшных» стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму, и устьичная щель открывается. При снижении интенсивности фотосинтеза (например, вечером) уменьшается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока в замыкающих клетках снижается по сравнению с околоустьичными. Вода путем осмоса уходит из замыкающих клеток, понижая их тургор, в результате ночью устьичная щель закрывается.
Клетки эпидермы плотно сомкнуты между собой, благодаря этому эпидерма выполняет целый ряд функций:
• препятствует проникновению болезнетворных организмов внутрь растения;
• защищает внутренние ткани от механических повреждений;
• регулирует газообмен и транспирацию;
• через нее выделяются вода, соли;
• может функционировать как всасывающая ткань;
принимает участие в синтезе различных веществ, восприятии раздражений и в движении листьев.
Трихомы — различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Их подразделяют на кроющие и железистые.Железистые трихомы, в отличие от кроющих, имеют клетки, выделяющие секрет.Кроющие волоски, образуя на растении шерстистый, войлочный или иной покров, отражают часть солнечных лучей и тем самым уменьшают транспирацию. Иногда волоски находятся только там, где расположены устьица, например, на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом значительно варьируют. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одно или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать, заполняясь воздухом.
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермальных слоев.
Анатомо-диагностические признаки, имеющие наибольшее значение и высокую вариабельность при определении лекарственного сырья. Волоски могут быть простые и головчатые, которые в свою очередь могут быть одноклеточными и многоклеточными. Многоклеточные волоски могут быть однорядными, двухрядными и ветвистыми.
Источник
2. Строение устьичных аппаратов.
Он состоит из двух замыкающих клеток (у двудольных бобовидной формы, у однодольных гантелеподобные). Между ними располагается устьичная щель, а под замыкающими клетками располагается подустьичная щель. Замыкающие клетки содержат хлоропласты и способны к фотосинтезу. В состав устьичного аппарата входят также побочные клетки.
Замыкающие клетки утолщены неравномерно: у двудольных растений те части, которые обращены к устичьной щели более толстые, а противоположные клеточной стенке – тонкие. У однодольных растений утолщённые клеточные стенки расположены на сторонах клеток, которые не прилегают к устичьной щели. Благодаря неравномерному утолщению клеточных стенок замыкающих клеток при повышении оводнённости замыкающих клеток они расходятся, открывая устичьную щель.
Побочные клетки (от 2 до 10) устьичных аппаратов могут иметь различную форму и разное строение. В зависимости от количества и расположения побочных клеток вокруг замыкающих клеток выделяют 6 типов устьичных аппаратов.
1. Аномоцитный тип – побочные клетки по размерам и форме не отличаются от остальных клеток эпидермы. Обычен для всех групп растений, исключая хвощи.
2. Диацитный тип – есть только две побочные клетки, общая стенка которых перпендикулярна устьичной щели (большинство губоцветных и гвоздичных растений).
3. Парацитный тип – побочные клетки располагаются параллельно замыкающим и устьичной щели (хвощи, папоротники и ряд цветковых растений).
4. Анизоцитный тип – замыкающие клетки окружены тремя побочными клетками, одна из которых заметно крупнее или мельче остальных (только у цветковых).
5. Тетрацитный тип – замыкающие клетки окружены четырьмя побочными клетками, из которых две латеральные, а две полярные (чаще у однодольных).
6. Энциклоцитный тип – побочные клетки образуют кольцо вокруг замыкающих клеток (папоротники, голосеменные, ряд цветковых).
Основная функция устьичных аппаратов – газообмен (поступление в лист СО2) и транспирация.
У растений, обитающих в условиях более или менее достаточного увлажнения (мезофиты), устьица расположены на нижней эпидерме, на верхней эпидерме их практически нет. Это помогает экономно расходовать воду. К ним относятся листопадные растения, луговые и лесные травы.
У растений, обитающих в условиях избыточного увлажнения (гигрофитов) устьица имеются на обеих эпидермах. К ним относятся кислица обыкновенная, папоротник орляк, щитовник мужской.
У растений, погружённых в воду только нижними частями (гидрофиты) с плавающими листьями устьица располагаются только на верхней эпидерме. К ним относятся кувшинка чисто белая, кубышка, стрелолист.
Когда растение начинает испытывать водный недостаток и содержание воды в листьях снижается ещё только на 2% (от 88 до 86%), содержание абсцизовой кислоты возрастает в 20 и даже 40 раз. Это вызывает закрывание устьиц, и потому потеря воды листьями уменьшается, растение меньше страдает при небольшой засухе. То же происходит при резком похолодании.
Из поглощенной корнями воды лишь 0,2% идёт на построение органики, а 99,8% испаряется.
Источник