8. Транспорт веществ по растению
Различают ближний и дальний транспорт веществ по растению. Ближний транспорт – это передвижение ионов, метаболитов и воды между клетками по симпласту и апопласту. Дальний транспорт – передвижение веществ между органами в растении по проводящим пучкам и включает транспорт воды и ионов по ксилеме (восходящий ток от корней к органам побега) и транспорт метаболитов по флоэме (нисходящий и восходящий потоки от листьев к зонам потребления веществ или отложения их в запас).
Загрузка сосудов ксилемы наиболее интенсивно происходит в зоне корневых волосков. В паренхимных клетках проводящего пучка, примыкающих к трахеидам или сосудам, функционируют насосы, выделяющие ионы, которые через поры в стенках сосудов попадают в их полости. В сосудах результате накопления ионов увеличивается сосущая сила, которая притягивает воду. В сосудах развивается гидростатическое давление и происходит подача жидкости в надземные органы.
Разгрузка ксилемы, то есть выход воды и ионов через поры сосудов ксилемы в клеточные стенки и в цитоплазму клеток мезофилла листа или клеток обкладки, обусловлена гидростатическим давлением в сосудах, работой насосов в плазмалемме клеток и влиянием транспирации, повышающей сосущую силу клеток листа.
Ассимиляты из клеток листьев поступают во флоэму, состоящую из нескольких типов клеток. В ситовидных трубках флоэмы плазмалемма окружает протопласт, содержащий небольшое число митохондрий и пластид, а также агранулярный эндоплазматический ретикулум. Тонопласт разрушен. Зрелая ситовидная трубка лишена ядра. Поперечные клеточные стенки – ситовидные пластинки – имеют перфорации, выстланные плазмалеммой и заполненные полисахаридом каллозой и фибриллами актиноподобного Ф-белка, которые ориентированы продольно. Ситовидные трубки связаны с клетками-спутниками плазмодесмами. Клетки-спутники (сопровождающие клетки) – это небольшие вытянутые вдоль ситовидных клеток паренхимные клетки с крупными ядрами, цитоплазмой, с большим количеством рибосом, других органелл и, особенно, митохондрий. Число плазмодесм в этих клетках в 3-10 раз больше, чем в стенках соседних мезофильных клеток. В клеточных стенках клеток-спутников много инвагинаций, выстланных плазмалеммой, что значительно увеличивает ее поверхность. Самые мелкие проводящие пучки включают один-два ксилемных сосуда и одну ситовидную трубку с сопровождающей клеткой. У многих С4-растений проводящие элементы листа окружены плотно сомкнутыми клетками обкладки, отделяющими пучки от мезофилла и от межклетников. Проводящая система листа представлена проводящими пучками, которые объединены в жилки разных размеров. Жилки расположены по листу так, чтобы обеспечить равномерный сбор ассимилятов по всей площади листа. Транспорт ассимилятов в листе строго ориентирован: ассимиляты передвигаются из каждой микрозоны клеток мезофилла радиусом 70-130 мкм в сторону ближайшего к ней малого пучка и далее по клеткам флоэмы в более крупную жилку.
Основной транспортной формой ассимилятов у большинства растений является сахароза (до 85 % от общего сухого вещества). Активность инвертазы – фермента, расщепляющего сахарозу на глюкозу и фруктозу – в проводящих тканях очень низка. Также транспортируются олигосахара, азотистые вещества, органические кислоты, витамины, гормоны. Неорганические соли составляют 1-3 % от общего количества веществ сока, особенно много ионов калия.
В клетках мезофилла осмотическое давление ниже, чем в тонких проводящих пучках. По мере продвижения от тонких пучков к средней жилке содержание сахаров возрастает. Поэтому загрузка проводящей системы ассимилятами идет против градиента концентрации с затратой энергии. Источником АТФ служат клетки-спутники. В плазмалемме клеток-спутников функционирует протонная помпа, выводящая наружу протоны. Она активируется ауксином и блокируется абсцизовой кислотой. Закисление апопласта в результате работы этой помпы способствует отдаче ионов калия и сахарозы клетками листа и поступлению их в клетки флоэмных окончаний. Трансмембранный перенос протонов происходит по концентрационному градиенту, а сахарозы – против градиента с помощью белков-переносчиков. Поступившие в клетки протоны вновь выкачиваются протонной помпой, работа которой сопряжена с поглощением ионов калия. Сахароза и ионы калия по плазмодесмам переносятся в полости ситовидных трубок.
В 1926 г. Э. Мюнх предложил гипотезу тока ассимилятов по ситовидным элементам флоэмы под давлением. Согласно этой гипотезе между фотосинтезирующими клетками листа, где накапливается сахароза, и тканями, использующими ассимиляты, создается осмотический градиент и возникает ток жидкости во флоэме от донора к акцептору. Предполагается также, что движущей силой перемещения жидкости из одной ситовидной трубки в другую через поры в ситовидной пластинке может быть транспорт ионов калия. Ионы калия активно входят в ситовидную трубку выше ситовидной пластинки, проникают через нее в нижележащую ситовидную трубку и пассивно выходят из нее в апопласт. В результате на ситовидных пластинках возникает электрический потенциал, способствующий транспорту веществ. Кроме того, фибриллы актиноподобного Ф-белка в порах ситовидных пластинок обладают сократительными свойствами и периодическими сокращениями способствуют передвижению жидкости по флоэме.
Разгрузка флоэмы происходит из-за высокого гидростатического давления в ситовидных трубках и аттрагирующей (притягивающей) способности органа-акцептора. Его аттрагирующая способность зависит от интенсивности роста органа, в ходе которого используются транспортируемые ассимиляты и тем самым снижается их концентрация в клетке. Следовательно, возникает градиент концентрации между элементом проводящей системы и клеткой акцептора. Интенсивность роста контролируется балансом регуляторов роста. В плазмалемме клеток акцептора функционирует протонная помпа, которая воздействует на ситовидные трубки и клетки-спутники, закисляя апопласт и тем самым способствует отдаче ими ионов калия и сахарозы в клеточные стенки. Затем сахароза поглощается клетками акцептора с участием мембранных переносчиков в симпорте с протонами, а ионы калия – по электрическому градиенту.
Источник
Мембранный транспорт ионов в растениях
Основной движущей силой поглотительной активности корней является работа ионных насосов. Ионы, поглощенные вместе с водой поступают в трахеиды и сосуды и транспортируются в остальные органы растения. В другие части растения ксилемный сок перемещается за счет транспирации и корневого давления. В клетках содержание тех или иных ионов бывает в десятки и сотни раз выше, чем в окружающей среде. Это связано с тем, что они способны к активному поглощению их против градиента концентрации и избирательному накоплению. Клеточные стенки, содержащие пектиновые вещества, благодаря которым она проявляет свойства катионообменников, т.е. способна быстро адсорбировать катионы. Далее, адсорбированные катионы проникают в клетку через мембраны. Растительные клетки отделены от внешней среды плазматической мембраной, через которую клетка получает необходимые элементы питания. Внутри клетки происходит разделение метаболизма по отдельным органеллам системой эндомембран. Во всех этих процессах очень важную роль играют различные мембранные переносчики ионов и органических молекул.
Транспорт заряженных частиц определяется электрохимическим потенциалом мембран. Цитоплазма живых клеток заряжена отрицательно по отношению к наружной среде. Т.е. существует разность электрических потенциалов, которая может служить движущей силой ионного транспорта. При этом облегчается поступление катионов внутрь клетки и выход из нее анионов.
Обычно концентрации, а, следовательно, химические потенциалы в разных частях клетки неодинаковы. Кроме того на мембранах поддерживаются градиенты электрических потенциалов. Это является причиной пассивного транспорта ионов в сторону более низкого электрохимического потенциала. Передвижение ионов по градиенту электрохимического потенциала называют пассивным транспортом, против градиента – активным. Различают следующие виды мембранного транспорта ионов:
3. диффузия через ионные каналы,
4. первично-активный транспорт,
5. вторично-активный (сопряженный) транспорт.
Пассивная диффузия ионов происходит при появлении в мембране различных гидрофильных пор или при нарушении целостности липидного бислоя. Липофильные вещества могут транспортироваться путем простой диффузии.
Облегченная диффузия осуществляется специализированными белками-переносчиками. В основе лежит обратимое связывание транспортируемого иона с белком-переносчиком, который проходит через мембрану с высвобождением иона на другой стороне. Облегченная диффузия характеризуется насыщением (когда все молекулы переносчика заняты ионами), селективностью и возможностью ингибирования.
Диффузия через селективные ионные каналы осуществляется по градиенту их электрохимического потенциала. Ионные каналы формируются интегральными белками, которые пронизывают мембрану таким образом, что в ней образуется гидрофильная пора. При этом гидрофильные аминокислоты выстилают стенки поры, а гидрофобные – контактируют с липидной фазой мембран. Движение ионов осуществляется в один ряд. В отличие от облегченной диффузии, транспорт ионов через каналы представляет процесс, который идет без насыщения и с более высокой скоростью. Кроме того, ионные каналы односторонни и селективны, что обусловлено диаметром просвета канала.
Первично-активный транспорт осуществляется ионными насосами, источником энергии для которых служит АТР, пирофосфат или субстраты, окисляемые в ЭТЦ митохондрий, хлоропластов. Для растительной клетки наибольшее значение имеют протонные и кальциевые АТРазы
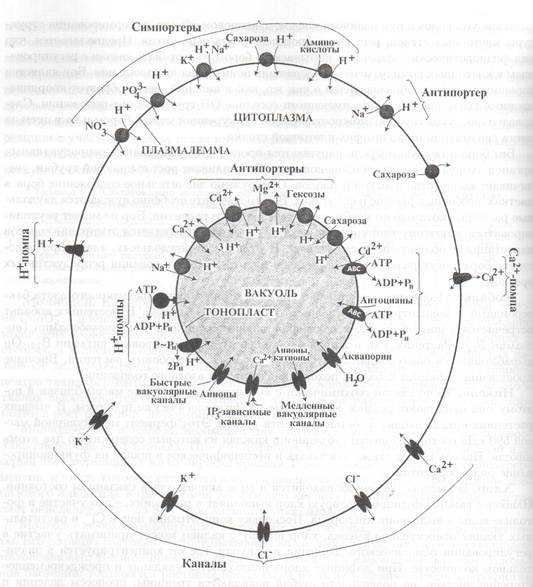
Вторично-активным (сопряженным) транспортом называют перенос ионов через мембрану против градиента его концентрации за счет энергии электро-химического градиента других ионов. Сопряженный транспорт может осуществляться в режиме симпорта (оба иона переносятся через мембрану в одном направлении) или антипорта (ионы транспортируются в разных направлениях) рис.. Для этой цели мембранные белки-переносчики
Рис. Система мембранного ионного транспорта на плазмалемме и тонопласте растительной клетки. Стрелками показано обычное направление ионных потоков.
чаще всего используют электрохимический градиент ионов водорода, создаваемый различными Н + -насосами.
Возможен транспорт веществ через мембраны путем экзоцитоза и пиноцитоза с образованием везикул.
Поступившие в клетку ионы далее транспортируются по корню растения по симпласту и апопласту. По апопласту (по межклетникам) перемещение ионов осуществляется за счет диффузии и ионной адсорбции по градиенту концентраций и ускоряется током воды. По симпласту (по цитоплазме) перемещение осуществляется благодаря движению цитоплазмы и по каналам ЭПС между клетками по плазмодесмам. Направление движения определяется градиентом ионов, который возникает из-за того, что поступившие в клетки ионы вовлекаются в обмен веществ.
Источник