Центры происхождения культурных растений
Значительный вклад в селекцию растений внес генетик Н. И. Вавилов. Он предположил и доказал, что все ныне выращиваемые в регионах мира культурные растения имеют свои определенные географические центры происхождения.
Не могу утаить от вас, что против блестящего ученого, Николая Ивановича Вавилова, было сфабриковано уголовное дело в 1940 году. Его великие труды были названы «вредительством», дело и любовь всей его жизни — «вражеской работой». Истинный гений умер в 1943 году в саратовской тюрьме, в состоянии дистрофии — жуткого истощения, из-за которого остановилось его сердце.

Вавилов выделил 7 основных географических центров происхождения культурных растений, которые расположены в поясах тропического и субтропического климата, где зарождалось земледелие. Продолжая исследования Вавилова, ученые открыли еще несколько подобных центров.
Итак, приступим к изучению центров происхождения и культурных растений, в них появившихся. Среди центров происхождения выделяют:
Лимонник, апельсин, соя, рис, просо, хурма, чайное дерево, редька, тутовое дерево (шелковица), бамбук, грецкий орех, женьшень.

Помело, бергамот, лайм, сахарная пальма, саговая пальма, мускатный орех, черный перец, гвоздичное дерево. Сахарный тростник — совместно с Индостанским центром.

Огурец, баклажан, фасоль, манго, базилик, мак, гречиха. Совместно с Индо-малайским центром — сахарный тростник, сахарная пальма.

Лук репчатый, дыня, чеснок, рожь, конопля, миндаль.

Рожь, пшеница, лен, горох, фундук, алыча, кизил, каштан, фисташка, виноград, айва, шпинат, эстрагон, финиковая пальма, лук-порей, барбарис.

Виноград, люпин, оливковое дерево, овес, капуста, брокколи, горчица, свекла, петрушка, сельдерей, редька, редис, брюква, спаржа, мята, хрен, укроп, щавель, мелисса.

Арбуз, кофе, сорго, кунжут, клещевина, просо, масличная пальма, кола.

Фасоль, кукуруза, батат, тыква, какао, авокадо, перец овощной, табак, подсолнечник.

Картофель, томат, хинное дерево, арахис, фейхоа, тыква мускатная, ананас, настурция.

Гомологические ряды в наследственной изменчивости
Это понятие введено Вавиловым Н.И. в результате его исследований, посвященных параллелизмам в явлении наследственной изменчивости. Внимательно изучите следующую цитату:
«Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов».
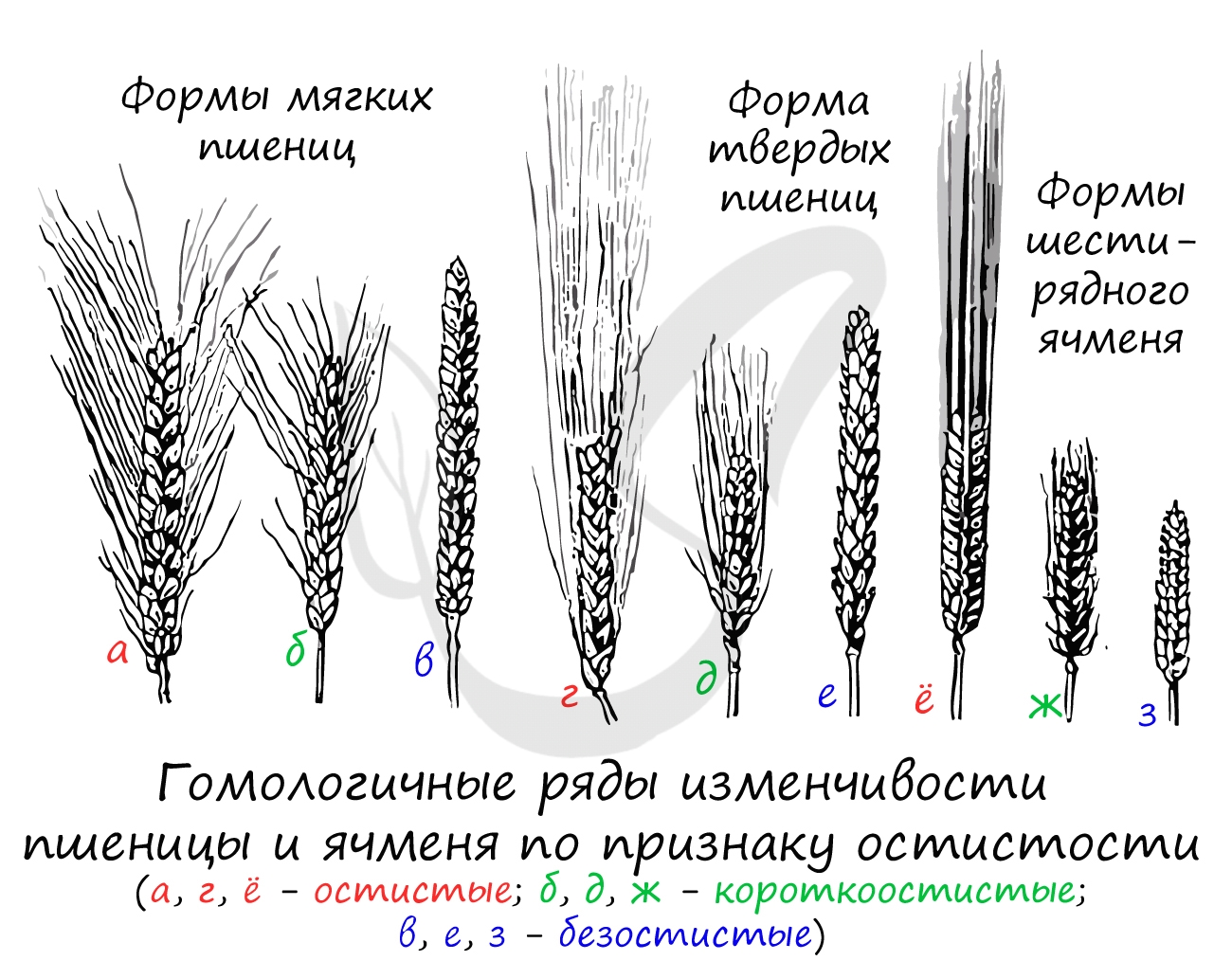
Суть явления кроется в сходных аллелях, повторяющихся у близких групп растений (и животных!), которые были обнаружены в ходе изучения наследственной изменчивости. Появилась возможность предсказывать еще не обнаруженные аллели у близкородственных групп растений (по одному «родственнику» судить о «другом»), что дало еще один импульс развитию селекции.
Важно и следующее следствие этого явления: зная совокупность мутаций, которые произошли в пределах одного вида, становится возможно предполагать их появление у другого, близкородственного вида.

© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Центры происхождения культурных растений (Таблица)
Таблица содержит центры происхождения основных наиболее значимых культурных растений.
Центры происхождения растений
Средиземноморье (Италия, Испания, Греция)
Маслина, свекла, редька, репа, капуста, лук, укроп, петрушка, мята, конопля, лен
Мягкая пшеница, рожь, ячмень, овес, бобовые, виноград, яблоня, груша, вишня, гранат, хурма, дыня, роза
Афганистан и Средняя Азия
Горох, чечевица, тыква, шпинат, морковь, репчатый лук, персик, абрикос, грецкий орех, миндаль
Восточно- Азиатский (Китайский)
Восточный и Южный Китай, Корея
Просо, ячмень, гаолян, гречиха, соя, киви, корица, чай
Рис, баклажан, огурец, манго, лимон, апельсин, сахарный тростник, джут, коротковолокнистый хлопчатник
Ямс. банан, кокосовая пальма. хлебное дерево, черный перец, мускатный орех
Твердая пшеница, сорго, горчица, кунжут, кофейное дерево
Мексиканский (Центрально- американский)
Мексика и Центральная Америка
Кукуруза (маис), томаты, батат, подсолнечник, стручковый перец, длинноволокнистый хлопчатник, какао, табак
Картофель, маниок, фасоль, ананас, арахис, гевея
Картофель, садовая земляника.
Южная Бразилия и Парагвай
Маниок, арахис, ананас, какао, гевея

Поделитесь ссылкой с друзьями:
Комментарии:
infotables.ru © 2014 — 2023. Копирование материала с сайта возможно только при наличие активной индексируемой ссылки на infotables.ru
Информация, размещенная на сайте, предоставлена в целях ознакомления. Владельцы сайта infotables.ru не несут ответственности за риски, связанные с использованием информации, полученной с этого cайта.
Источник
Центры происхождения культурных растений (по н. И. Вавилову).
Тропическая Индия, Индокитай, Южный Китай, о-ва Юго-Вос-точной Азии.
Рис, сахарный тростник, цитрусовые, огурец, баклажан, чер-ный перец и др. (50 % культурных растений).
Центральный и Восточный Китай, Япония, Корея, Тай-вань.
Соя, просо, гречиха, плодовые и овощные культуры – слива, вишня, редька и др. (20 % культурных рас-тений).
Малая Азия, Средняя Азия, Иран, Афганис-тан, Юго-Западная Индия.
Пшеница, рожь, бобо-вые культуры, лен, конопля, репа, мор-ковь, чеснок, виноград, абрикос, груша и др. (14 % культурных растений).
Страны по берегам Средиземного моря.
Капуста, сахарная свекла, маслины, кле-вер, чечевица, кормо-вые травы (11 % культурных растений).
Абиссинское нагорье Африки.
Твердая пшеница, ячмень, кофейное де-рево, сорго, бананы.
Кукуруза, длинново-локнистый хлопчатник, какао, тыква, табак.
Южная Америка вдоль западного побережья.
Картофель, ананас, хинное дерево.
Таким образом, обнаружение спонтанных или индуцированных мутаций у одного вида дает основания для поисков сходных мутаций у родственных видов растений или животных.
Закон гомологических рядов наследственной изменчивости успешно используется в селекционной практике. Работа по созданию семенных коллекций сортов культурных растений и их дикорастущих предков, начало которой положил Н. И. Вавилов, в наше время продолжается. В России в настоящее время коллекция включает более 320 тыс. образцов, относящихся к 1041 виду растений. Сюда входят дикие виды, сородичи культурных растений, старые местные сорта, все лучшее и новое, что создано за последнее время усилиями селекционеров всех стран мира. Из мирового генофонда ученые выделяют генетические источники хозяйственно ценных признаков: урожайности, скороспелости, устойчивости к болезням и вредителям, засухоустойчивости, устойчивости к полеганию и др. Современные генетические методы дают возможность добиваться в селекции растений очень крупных успехов. Так, использование ценных генов дикого эфиопского ячменя позволило создать выдающийся по продуктивности сорт ярового ячменя Одесский 100.
№ 70. Особенности наследования хозяйственно-полезных признаков.
Большинство признаков и свойств организмов характеризуются количественным типом индивидуальной изменчивости, для которой типично непрерывное изменение величины признака у особей какой-либо группы. Величина количественного признака варьирует от минимального уровня у части особей к среднему – у других и далее к максимальному уровню у остальных. Даже в пределах достаточно однородной по полу, возрасту, породе группы животных у близкородственных особей наблюдается индивидуальная изменчивость признака, величину которого можно измерить. К количественным признакам относят хозяйственно ценные (живая масса, величина, удой, настриг шерсти) и физиологические признаки. Они характеризуются типичным непрерывным изменением уровня у особей конкретной группы. К количественным признакам относят также и те, которые имеют прерывистое выражение, например яйценоскость, плодовитость, а также ряд физиологических отличий.
Количественные признаки непрерывного и прерывистого типов изменчивости имеют важное значение в практике животноводства и ветеринарии, в научных исследованиях.
Наследование количественных признаков обусловлено одинаковым или сходным действием многих доминантных неаллельных генов на признак (полимерия) либо многими однозначными генами (полигения). На наличие двух или трех пар однозначно действующих полимерных генов, определяющих степень выраженности признака, указывает тип расщепления признака у особей второго поколения. Так, при трех доминантных генах А1; А2 и А3 и их рецессивных аллелях а1, а2, а3 во II поколении будут выявлены 64 варианта генотипов в соотношении 1:6:15:20:15:6:1.
Если общая возможность развития признака связана с действием одного гена, то его принято называть главным (менделирующим) геном (олигогеном) и тогда признак наследуется в соответствии с законом Менделя. Полигены могут проявлять модифицирующее влияние на количественные признаки и составлять группу генов-модификаторов, то есть генов, которые, действуя каждый отдельно, проявляют слабое влияние на изменение в фенотипе, вызванное действием главного гена. Гены-модификаторы могут оказывать влияние и при отсутствии главного гена.
Полимерные гены способствуют увеличению изменчивости и формированию различных подгрупп (экотипов) внутри вида, так как они обеспечивают многообразные рекомбинации генотипов. Влияние рекомбинации и отбора в разных условиях среды способствует образованию несходных экотипов и повышению приспособленности вида к многообразию факторов внешней среды, формированию наследственной адаптации.
За последнее десятилетие расширились представления о наследовании и изменчивости признаков. Ранее полагали, что особенности потомства обусловлены свободной перекомбинацией генов как в процессе гаметогенеза, так и при оплодотворении. Однако многочисленные исследования показали, что изменчивость потомства, определяемая действием неаллельных генов, вызывается не только возможностью их случайной перекомбинации, а обусловлена определенной комбинацией гамет родителей. Таким образом, создается источник изменчивости и наследственности потомства за счет комбинаторики несходных гамет родителей. Несходство гамет по генетическому материалу может иметь место как среди гамет самца, так и среди гамет самки. При комбинации несходных гамет родителей возникают новые межлокусные корреляции, происходит изменение частоты генов в данном локусе у потомства по сравнению с их частотой у исходных родительских форм. Создается новый тип связи между генетическими элементами у потомства, который получил название «гаметическая интеграция».
Гаметическая интеграция способствует формированию устойчивости разнообразных признаков и свойств, благоприятствующих повышению приспособленности, так как образуются адаптивные комплексы генов, которые повышают индивидуальную изменчивость организмов по степени их приспособленности к условиям среды. Гаметическая интеграция межлокусных корреляций обусловливает на фенотипическом уровне взаимодействие между локусами неаллельных полигенов. Она может быть выявлена при помощи статистических параметров.
Приспособленность организмов могут характеризовать величины среднего уровня жизнеспособности особей, их плодовитость, интенсивность развития всех особей данной группы (популяции).
Если в локусе одной из хромосом гаметы образовалась мутация, имеющая адаптивное значение, то при передаче ее в последующие поколения будет повышаться частота распространений мутации и одновременно на фоне гаметической интеграции будет увеличиваться частота других генов, входящих в хромосомы этой гаметы. Явление получило название «попутный транспорт генов».
Обобщение теории роли гаметической интеграции было сделано в трудах Майра, 1974; Левонтина, 1978; Животовского, 1984; и др.
Теоретическая работа Фальконера (1985) показала, что популяционная средняя величина признака характеризует не только фенотипический его уровень, но и генотипический уровень в ряде поколений при сохранении факторов среды. Изменение среднего уровня признака при отборе происходит более эффективно в сторону его увеличения, чем в сторону уменьшения, что могло быть результатом действия «сильных» генов и материнского эффекта. Выяснено, что чем большее число локусов определяет уровень признака и чем длительнее отбор в ряде поколений, тем более выражена реакция на отбор.
Возникает вопрос: существуют ли тупики отбора, когда его действие прекращается? Оказывается, что постоянный процесс мутирования создает новый источник изменчивости и тупика отбора не возникает.
Для выявления наследуемости признака широко применяют методы корреляционного и дисперсионного анализа. Тем самым выявляется доля генетического влияния на признак в фенотипической изменчивости.
Источник