Характеристика гаметофита и спорофита: определения и особенности
Гаметофит — это гаплоидная фаза жизненного цикла высших растений и водорослей, которая берет свое начало из спор и производит, в результате, половые клетки или гаметы.
Спорофит — это диплоидная фаза жизненного цикла растений и водорослей, результатом которой является производство спор.
В жизненном цикле растений наблюдается последовательное чередование спорофита и гаметофита. Гаметофиту отводится роль реализации полового размножения, а спорофиту — обеспечение бесполого типа воспроизведения организма.
У водорослей спорофит возникает из зиготы — внутри него формируются споры. Зигота образуется на половой гаплоидной фазе из женской гаметы — яйцеклетки. Важно, чтобы яйцеклетку оплодотворила мужская половая клетка.
Гаметофит — это гаплоидная фаза в жизненном цикле растительного организма. Он получает развитие из спор и формирует женские и мужские половые клетки. В результате слияния гамет формируется зигота — она дает гаплоидное поколение.
Формирование мужских гамет осуществляется из гаметангий или антеридий. У споровых и семенных растений гаметы имеют разные названия: в первом случае — сперматозоиды, во втором — спермиями.
Сперматозоиды подвижны — эта подвижность достигается за счет жгутиков. Женские гаметы, для сравнения — это неподвижная яйцеклетка: их образование происходит внутри женских гаметангиев.
Архегонии являются женскими гаметангиями растений.
Оплодотворение яйцеклетки наземной растительности осуществляется внутри архегония. Затем происходит спорообразование, и жизненный цикл проходит собственное чередование.
Наиболее заметно чередование поколений у споровых растений. Наблюдается раздельное нахождение гаметофита и спорофита у таких групп растений как:
У этих групп растений преимущество получило гаплоидное поколение. Жизненный цикл папоротников — хороший пример чередования поколений. Гаметофит у них представлен в виде маленького и недолговечного ростка, на котором вырастает спорофит.
У мхов фазы не разделяются — в этом случае коробочка со спорами формируется на гаметофите. Что касается цветковых растений, то мужские гаметы здесь формируются внутри тычинок, которые осуществляют перенос мужских половых клеток на пестики.
Гаметофит голосеменных имеет особенность. У голосеменных растений семена находятся в открытом виде в шишках, при этом процесс оплодотворения практически не отличается от процесса, происходящего у покрытосеменных. Развитие семени голосеменных происходит из семязачатка (он открыт на семенной чешуе), а у покрытосеменных семя находится внутри плода (мужской гаметофит покрытосеменных).
Выделяют несколько этапов в жизненном цикле растительных организмов:
- формирование спорофита и спор;
- преобразование спорофита и спор в молодой гаметофит;
- образование гамет.
Особенности гаметофита и спорофита
Остановимся подробнее на том, что такое спорофит и гаметофит.
Гаметофит и спорофит растений отличаются как минимум по двум критериям: размеру и этапу оформления.
Также, к примеру, гаметофит относится к гаплоидам и характеризуется наличием одинарного набора хромосом. Спорофит же относится к диплоидам и обладает двойным набором хромосом.
Гаметофит обеспечивает половое размножение, а спорофит — бесполое.
С началом в гаметофите мейоза происходит запуск процесса образования гамет — эти гаметы отличаются высокой активностью.
У большинства растений в жизненном цикле преобладает спорофит. Это связано с определенными преимуществами, которые он обеспечивает. Если среда водная, то у гамет есть возможность передвигаться. А вот в наземно-воздушной среде у растений нет возможностей для перемещения собственных спор. Диплоидные растения легче сохраняют рецессивные признаки в сложных условиях на поверхности земли. Эти признаки в дальнейшем могут обеспечить выживание.
У разных групп растений соотношение спорофита и гаметофита разное. К примеру, для высших растений (за исключением мхов) характерно преобладание гаплоидности. Этому есть объяснение: в природе наличие семени важно для дальнейшей жизни. Гаметофит отвечает за реализацию непосредственного оплодотворения. Споры же нужны для распространения и произрастания вида на планете.
Только диплоидные организмы способны справляться с резкой сменой условий обитания. По этой причине диплоидное поколение преобладает у наземных растений, а у подводных растений чаще встречается гаметофит или гаплоидная часть.
К примеру, для одноклеточных водорослей (хламидомонад) характерно преобладание гаплоидности на протяжении всего периода жизни.
Отличия гаметофита от спорофита заключаются в:
- мейоз — способ образования спор из спорофита;
- гаметофит образуется из споры;
- как результат мейоза — образование половых клеток из гаметофита.
Для зиготы и спорофита характерны диплоидные свойства, а вот споры и гаметофит связаны с гаплоидностью.
Спорофит мхов (прорастание споры) начинается с «зеленой нити», затем предросток дает начало гаметофиту. Споры семенных растений прорастают непосредственно внутри спорофита. Макроспора прорастает в женский гаметофит — в нем происходит образование яйцеклетки. Мужской гаметофит прорастает в пыльцевое зерно — в нем образуются спермии.
В изучении особенностей спорофита и гаметофита важны следующие моменты:
- гаметофит папоротников, гаметофит плаунов и хвощей — это их заросток;
- образование половых клеток происходит только в органах гаметофита;
- женский гаметофит цветковых растений — это зародышевый мешок. В нем находится 7 клеток, в том числе яйцеклетка и центральная клетка с диплоидным набором хромосом;
- в состав мужского гаметофита входят вегетативная клетка, превращающаяся в пыльцевую трубку, и генеративная клетка, продуцирующая образование двух спермиев. Один из спермиев сливается с яйцеклеткой, а второй участвует в оплодотворении центральной клетки. Как результат — появляется диплоидная зигота и триплоидный эндосперм.
Разделение спорофита и гаметофита — важный этап эволюционного процесса растений, который обусловил их высокий уровень приспособленности к окружающей среде.
Источник
Мегаспорогенез и развитие женского гаметофита
Наиболее важную роль в семязачатке играет нуцеллус, или мегаспорангий, в котором образуются мегаспоры и впоследствии развивается женский гаметофит.
Как уже было сказано, еще на ранней стадии развития семязачатка в нуцеллусе обособляется единственная археспориальная клетка (очень редко возникают 2 и более клеток), непосредственно превращающаяся в мегаспо-роцит. По размерам он больше остальных клеток нуцеллуса, имеет плотную цитоплазму и крупное диплоидное ядро. В результате редукционного деления ядра мегаспороцита возникают гаплоидные мегаспоры, составляющие линейную тетраду (рис. 244 В, 1, 3). Из самой нижней, сильно разрастающейся мегаспоры, в дальнейшем развивается женский гаметофит, а остальные мегаспоры отмирают (рис. 245 А).
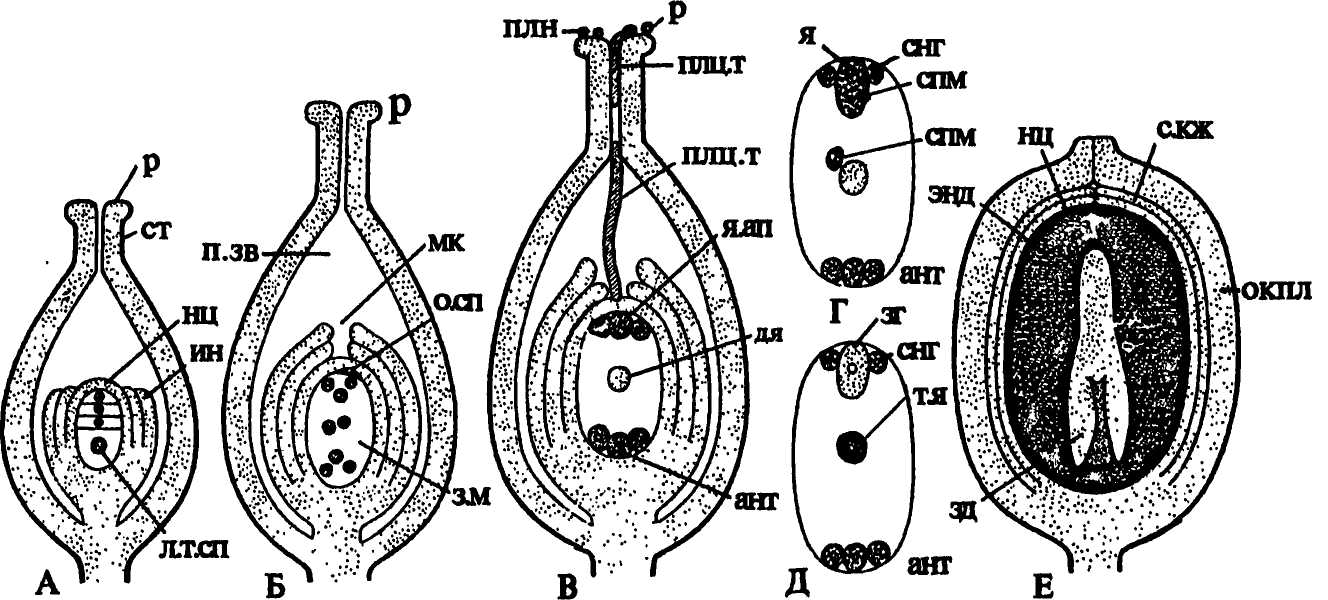
Рис. 245. Развитие зародышевого мешка, оплодотворение, строение односеменного плода: А — тетрада мегаспор; Б — 8-ядерный зародышевый мешок; В — дифференциация клеток внутри зародышевого мешка; Г — двойное оплодотворение; Д — образование зиготы и вторичного ядра зародышевого мешка; Е — односеменной плод в продольном разрезе.
Обозначения: ант — антиподы, д.я — диплоидное ядро, зг — зигота, зд — зародыш, з.м — зародышевый мешок, ин — интегументы, л.т.сп — линейная тетрада мегаспор, мк — микропиле, нц — нуцеллус, окпл — околоплодник, о.сп — 3 отмершие мегаспоры, п.зв — полость завязи, плн — пылинки, плц.т — пыльцевая трубка, р — рыльце, с.кж — семенная кожура, снг- синергиды, спм — спермий, ст — столбик, т.я — триплоидное ядро, энд — эндосперм, я — яйцеклетка, я.ап — ядерный аппарат
Женский гаметофит покрытосеменных называют зародышевым мешком. Он формируется следующим образом. Ядро увеличившейся в размерах мегаспоры митотически делится, возникшие 2 ядра расходятся к разным полюсам клетки. В ее центральной части формируется крупная вакуоль. При втором д елении образуются 4 ядра, при третьем — восемь. К началу клеточной дифференциации зародышевого мешка исходная клетка достигает значительных размеров (рис. 245 Б). Параллельно с ее ростом разрастается и весь семязачаток, нуцеллус, как правило, истончается, а иногда, как у сложноцветных, исчезает полностью, так как содержащиеся в нем питательные вещества расходуются на развитие заростка. От каждого полюса клетки в ее центральную часть отходит по одному ядру, которые впоследствии сливаются, образуя диплоидное ядро центральной клетки зародышевого мешка.
На микропилярном полюсе зародышевого мешка вокруг каждого из 3 оставшихся ядер обособляется цитоплазма, формируются оболочки и образуется яйцевой аппарат, состоящий из яйцеклетки и двух вспомогательных клеток — синергид. Яйцеклетка обычно крупнее, ее ядро находится в нижней части, под вакуолью, а в синергидах, наоборот, ядра располагаются в верхних частях клеток, над вакуолями. На халазальном полюсе зародышевого мешка таким же путем возникают 3 антиподы (рис. 245 В, Г).
Описанный, наиболее часто встречающийся 8-ядерный зародышевый мешок называют моноспорическим. Он образуется из одной (самой внутренней) мегаспоры тетрады. Однако 8-ядерные зародышевые мешки могут формироваться и другими способами (рис.246). У лука после 1-го деления мейоза возникает диада клеток. Одна клетка дегенерирует. Ядро второй клетки делится митотически, образующиеся ядра расходятся к разным полюсам клетки. Каждое из этих ядер соответствует ядру одной из 2 мегаспор диады. Формирующийся из двуядерной клетки зародышевый мешок называют биспорическим. У адоксы мегаспорогенез вообще не сопровождается цитокинезом, и каждое из 4 образующихся ядер соответствует ядрам неразвившейся тетрады мегаспор. Возникший из такой 4-ядерной клетки зародышевый мешок называют тетраспорическим.
Эти примеры показывают, что у покрытосеменных растений существуют разные механизмы, ускоряющие развитие женского гаметофита. Другие отклонения в строении зародышевого мешка связаны с уменьшением или увеличением числа клеток и числа ядер, их плоидности. Особенно сильно варьируют числа антипод.
Таким образом, женский гаметофит покрытосеменных по сравнению с женскими гаметофитами всех других разноспоровых растений достиг крайней степени редукции. Он состоит из крупной диплоидной центральной клетки, окруженной оболочкой мегаспоры, и 6 находящихся внутри нее мелких клеток. Женский гаметофит покрытосеменных растений утратил архегонии, сохранив способность к формированию женской гаметы — яйцеклетки.
Естественно, возникает вопрос о возможности гомологизации зародышевого мешка с женскими заростками других разноспоровых растений. Все многочисленные точки зрения практиче ски могут быть сведены к 3 основным.
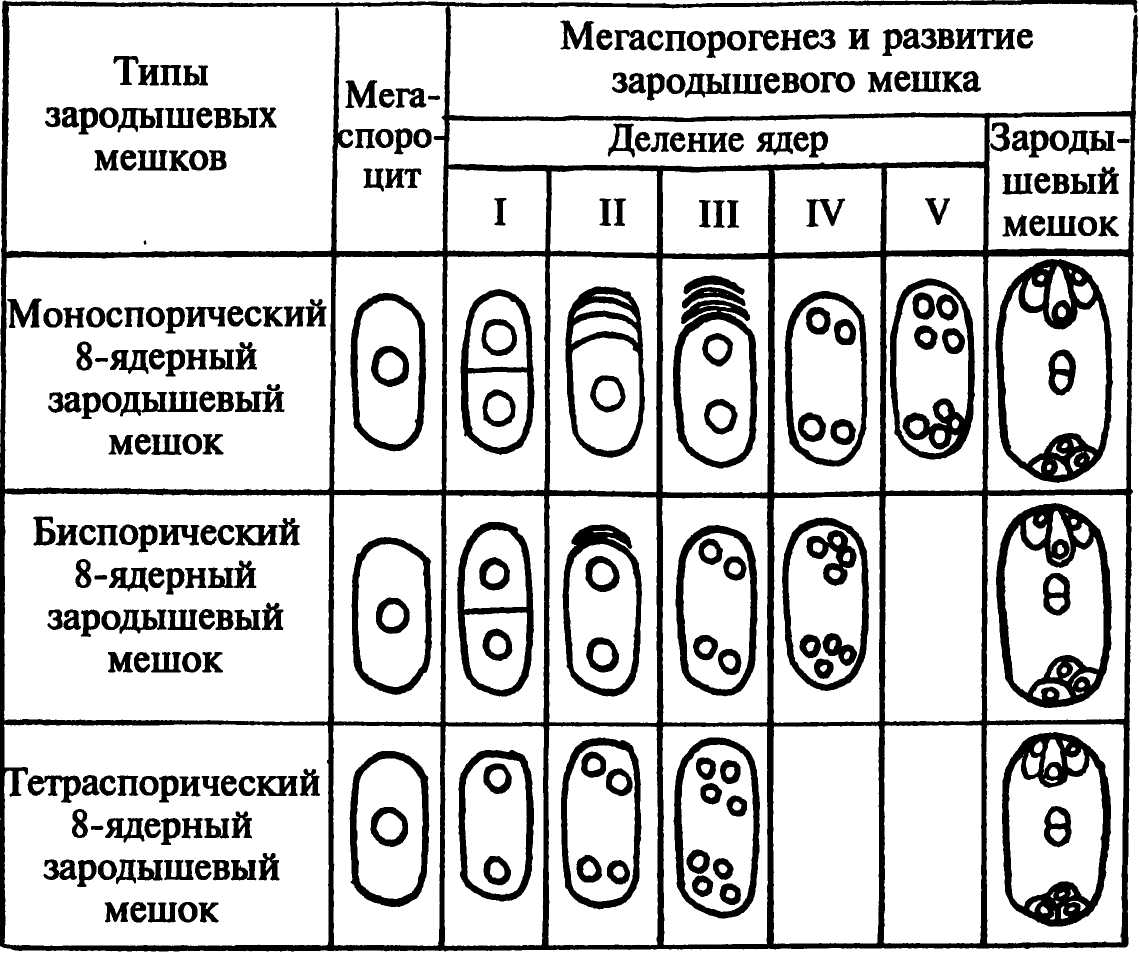
Рис. 246. Разные типы формирования 8-ядерных зародышевых мешков
- Зародышевый мешок — типичный гаметофит, состоящий из вегетативной части (синергид и антипод) и редуцированного архегония, от которого сохранилась одна яйцеклетка.
- Зародышевый мешок имеет 2 редуцированных архегония, расположенных в микропилярной и халазальной его частях. При этом яйцеклетку считают гомологом одной из антипод. 2 синергиды и 2 антиподы рассматривают как шейковые канальцевые клетки, а полярные ядра — как ядра брюшных канальцевых клеток. Однако в настоящее время выяснено, что синергиды представляют собой сестринские клетки, а ядро яйцеклетки — сестринское одному из полярных ядер. Таким образом, эта гипотеза оказалась несостоятельной.
- Зародышевый мешок — гомолог женского гаметофита остальных разноспоровых растений, так как возникает из мегаспоры, но он настолько видоизменился в процессе эволюции, что нецелесообразно пытаться найти гомологию между его отдельными структурами.
8- ядерный зародышевый мешок соответствует свободно-ядерной стадии развития женского гаметофита голосеменных и некоторых разноспоровых папоротников, например, сальвинии. При общности начальных стадий развития всех женских гаметофитов, конечные стадии развития зародышевого мешка сильно изменились и приобрели специфические особенности, не имеющие гомологов среди голосеменных. В связи с этим, формирование зародышевого мешка можно рассматривать как следствие эволюционного ускорения, сопровождающегося утратой способности к образованию типичных архегониев и вегетативной части заростка. Кстати, этот процесс, хотя и в другом варианте, происходил и у гнетовых из отдела голосеменных, у которых отмечены все стадии развития женских заростков — от нормально развитого эндосперма с архегониями у эфедры до своеобразных гаметофитов, не имеющих архегониев, у гнетума и вельвичии.
Источник