Отличительные характеристики и черты высших споровых растений
Отличительные характеристики высших споровых растений
Какие растения называют споровыми?
Споровые растения — это такие растения, размножающиеся спорами и распространенные в наземной среде.
Споровые растения представляют собой новый этап эволюционного развития растительного мира. Высшие растения — это растения, которые имеют расчленение тела на вегетативные органы: корень, стебель и листья. У низших растений такой дифференцировки нет. Признаком высших растений является еще и то, что вегетативные органы строятся из разнообразных тканей.
Высшие споровые растения обычно являются обитателями суходола. Однако встречаются и обитатели водоемов.
Все наземные листостебельные растения, размножающиеся спорами, относятся к высшим споровым растениям. Среди них представители отделов:
- Мохообразные или Мхи, представленные 25 тысячами видов;
- Плаунообразные или Плауны, которых насчитывается около 400 видов;
- Хвощеобразные или Хвощи. Это 32 вида;
- Папоротникообразные или Папоротники. Их примерно 10 тысяч видов.
Основные примеры споровых растений — различные виды папоротников, мхов, лишайников и др.
Высшие растения сформировались давно. Среди предков высших растений называют бурые или зеленые водоросли, достигшие высокого уровня дифференциации тела и органов полового размножения.
Низшие споровые растения, как правило, заселяют водную среду. На суше господствующее положение занимают высшие растения. С выходом на сушу у последних появились специальные приспособления к новым условиям существования.
Отличительные черты высших споровых растений
Каковы отличительные особенности высших растений по сравнению с низшими?
Растения, которые размножаются спорами, имеют следующие особенности:
- разные типы тканей. С внешней стороны растения имеют ткань, которая призвана защищать их от неблагоприятных условий. Процесс фотосинтеза обеспечивает отлично развитая хлорофилоносная ткань. Имеющиеся проводящие ткани обеспечивают протекание обмена веществ между подземными и надземными органами. Механические (опорные) и запасающие ткани тоже хорошо развиты;
- дифференцировка тела на органы. В первую очередь у высших растений произошло формирование специальных органов, отвечающих за поглощение минеральных веществ из субстрата. Это ризоиды и корневые волоски. Автотрофное питание — общая биологическая черта всех высших растений. По этой причине у них произошло формирование фотосинтезирующего органа — листка. Формирования стебля и корня произошло ввиду необходимости связи двух важных конечных аппаратов: корневого волоска и зеленой клетки листка. Также одна из причин — обеспечение стойкости растения в воздушной среде и почве;
- многоклеточные органы полового размножения. Они встречаются двух типов: мужские (антеридии) и женские (архегонии);
- начало онтогенеза происходит с зародыша, который развивается из зиготы. Она представляет собой клетку, образованную в результате слияния двух гамет;
- правильное чередование полового и бесполого поколений. Гаметофит — половое поколение, спорофит — бесполое;
- преобладание спорофита в цикле развития. Исключение — мохообразные (понятно, по каким признакам мхи относят к высшим растениям). Причина прогрессивного развития спорофита — высокая приспособленность к наземным условиям и большим объемом генетической информации (она свойственна диплоидному поколению).
Перечисленные особенности позволяют ответить на вопросы, почему мхи называют высшими споровыми растениями, по каким признакам мхи относятся к высшим растениям, а также почему папоротники относят к высшим растениям.
Жизненный цикл высших споровых растений и чередование поколений
Под жизненным циклом понимают последовательность фаз в развитии растения, по окончании которых растительный организм достигает зрелости и становится способным дать начало будущим поколениям: половому (гаметофиту) и бесполому (спорофиту).
У высших споровых растений жизненный цикл состоит из ритмического чередования двух поколений: бесполого и полового (спорофита и гаметофита соответственно).
На спорофите происходит образование спорангиев — органов бесполого размножения. В них происходит формирование спор. Из спор формируются гаметофиты, особи полового поколения, среди которых встречаются как одно-, так и двуполые.
На гаметофите происходит образование антеридий и архегоний — мужских и женских половых органов соответственно. В антеридиях формируются подвижные сперматозоиды, а в архегониях — неподвижные яйцеклетки.
Капельножидкая вода — ключевое условие для осуществления оплодотворения. Вода нужна для того, чтобы сперматозоид мог перемещаться к яйцеклетке. Оплодотворенная яйцеклетка дает начало зародышу, которые постепенно растет и превращается в особь бесполого поколения — спорофит.
Важно, что зигота дает начало только спорофиту, а спора — гаметофиту.
Спорофит преобладает в цикле развития большинства споровых растений, кроме мохообразных. Он имеет лучшую приспособленность к жизни в сложных условиях наземной среды. Из этого следует, что для эволюции высших споровых растений, за исключением мхов, свойственна тенденция к преобладанию и усовершенствованию спорофита (параллельно происходит редукция гаметофита).
Источник
Жизненные циклы высших растений.
У всех наземных растений, как и у наиболее высокоорганизованных водорослей, наблюдается чередование поколений. В жизненном цикле (т.е. цикла от зиготы одного поколения до зиготы следующего поколения) один тип организма сменяется другим. Гаплоидное поколение называется гаметофитом, так как оно способно к половому размножению и образует гаметы путем митоза. Сливаясь, гаметы образуют диплоидную зиготу, из которой вырастает следующее поколение – диплоидные спорофиты. Спорофит способен к бесполому размножению с образованием спор. Споры образуются путем мейоза, т.е. наблюдается возврат к гаплоидному состоянию. Гаплоидные споры дают начало гаметофитному поколению. Одно из двух поколений всегда преобладает над другим, на его долю приходится большая часть жизненного цикла (доминирующее поколение).
Эволюция растений.
Водоросли – низшие растения, их тело (слоевище) не разделено на органы, ткани отсутствуют. Они бывают одноклеточными, колониальными и многоклеточными. Размеры от микроскопических до нескольких десятков метров. Клеточная стенка из целлюлозы и пектиновых веществ сочетает опорную и защитную функцию с возможностью ростовых процессов и проницаемостью. В цитоплазме особенно заметны хроматофоры, которые различны по величине, форме, числу, местоположению, строению, набору пигментов. Размножение водорослей происходит бесполым (фрагментация, споры) и половым способом. У многих наблюдается чередование поколений в жизненном цикле.
Красные водоросли обычно морские, реже пресноводные. Чаще многоклеточные. Размеры от нескольких миллиметров до метра в длину. Внешне многообразны: нитевидные, цилиндрические, пластинчатые и кораллоподобные, в разной мере рассеченные и разветвленные. Самые глубоководные – в морях живут на глубине до 270 м, куда проникает зеленый, синий и фиолетовый свет, поэтому кроме хлорофилла содержат красные и синие пигменты. Представители: филлофора, порфира и др.
Бурые водоросли почти все морские; форма нитчатая и талломовидная, обычно крупные. Живут на небольшой глубине, часто в зоне приливов. Кроме хлорофилла содержат бурый пигмент. Запасают много углеводов и жиров. Используют в пищу, в косметологии, медицине. Представители: фукус, ламинария.
Зеленые водоросли очень разнообразны – одноклеточные, нитчатые, талломовидные и др. Морские и пресноводные, некоторые приспособились к жизни на суше. Кроме хроматофоров в клетках могут содержаться хлоропласты. Среди фотосинтетических пигментов преобладает хлорофилл. Запасают углеводы в виде крахмала. Хламидомонада, хлорелла (одноклеточные); спирогира, улотрикс (нитчатые); ульва (талломовидная)
Водоросли



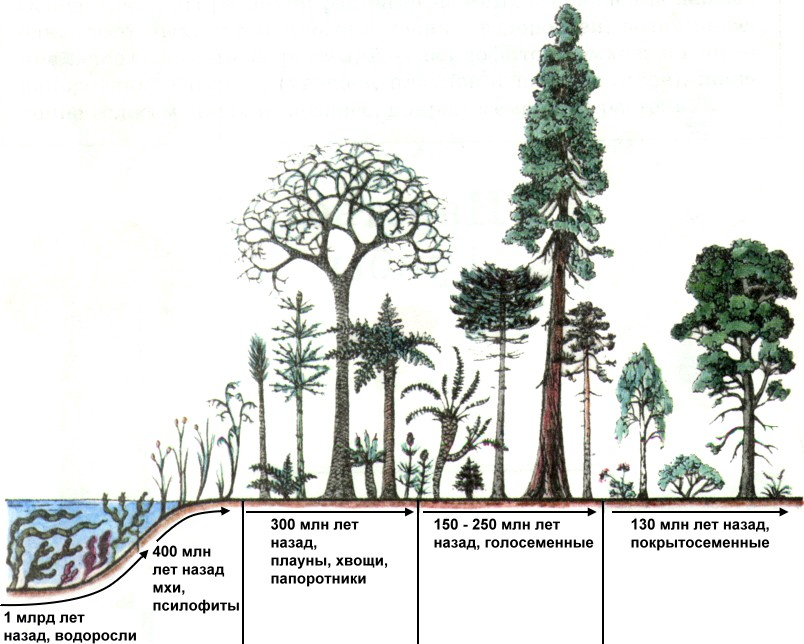
Выход на сушу. Псилофиты. Существует предположение, что высшие растения произошли от древних зеленых водорослей. Первые высшие растения появились на суше примерно 430 млн. лет назад в виде небольших по размеру и примитивных по строению риниофитов и псилофитов. Они еще не имели стеблей, листьев, корней, а представляли собой ветвящиеся оси, на подземных частях которых развивались ризоиды, а на концах надземных веточек развивались спорангии со спорами. От водорослей они отличались и более сложным строением: у них были развиты покровная, механическая и проводящие ткани.
У мхов нет корней (к почве крепятся ризоидами) и не развиты проводящие ткани. В жизненном цикле мхов преобладает половое поколение — гаметофит; большинство экземпляров раздельнополы; на мужских особях в антеридиях развиваются сперматозоиды; на женских особях в архегониях развиваются яйцеклетки; для оплодотворения нужна капельно-жидкая влага; после оплодотворения на женском гаметофите из зиготы развивается спорофит (коробочка), который полностью от него зависит и питается за его счет; в коробочке созревают споры (образуются путем мейоза); из споры развивается предросток – протонема (нитчатая или пластинчатая), на которой образуются почки, дающие начало новым гаметофитам.
К Моховидным относят печеночные мхи (риччия, маршанция) и листостебельные мхи (кукушкин лён, сфагнум).
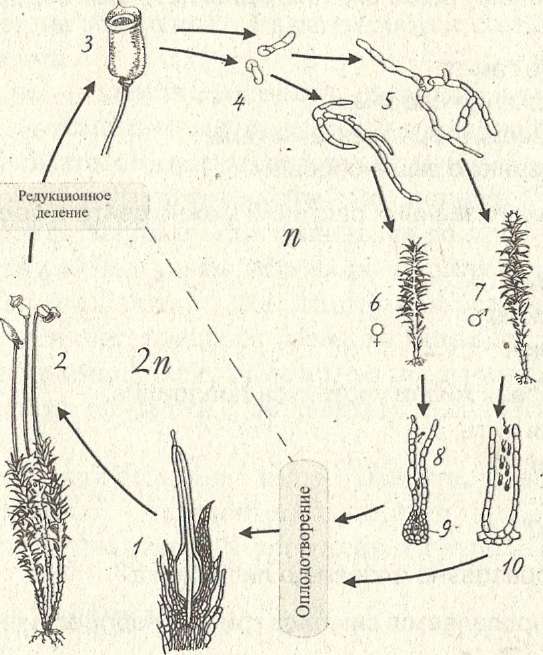



Высшие споровые растения. Чередование поколений, доминирует спорофит. У спорофита развиваются настоящие корни (придаточные), стебли, мелкие листья. Спорангии собраны в спороносные колоски – стробилы. Гаметофит обычно подземный, развивается медленно – до 10-15 лет. Всего около 1000 видов. Представители – плаун булавовидный, селагинелла.


Высшие споровые растения. Чередование поколений, доминирует спорофит. Мутовчатое расположение ветвей и листьев. Листья в виде пленчатых выростов в узлах. Функцию фотосинтеза несут побеги. Стебель и боковые веточки жесткие, ребристые, содержат кремнезем. Характерно длинное ветвящееся корневище. Споры развиваются в спорангиях, собранных в стробилы обычно на верхушках побегов. Гаметофит наземный, фотосинтезирующий, слоевищного типа, прикрепляется к почве ризоидами. Представители: хвощ полевой; хвощ зимующий.
Отдел Папоротниковидные.
В жизненном цикле доминирует спорофит. Главный корень очень рано отмирает и замещается системой придаточных корней, часто развивается корневище (ползучее или укороченное). Листья – вайи, соответствующие крупным системам ветвления (плосковетка). У большинства вайи перистые. Спорангии обычно собраны в мешочки — сорусы, расположенные на нижней части листа.
Споры разносятся ветром, при попадании в благоприятные условия прорастают в гаметофит — пластинчатый заросток, прикрепляющийся к субстрату ризоидами. На нижней стороне заростка в архегониях созревают яйцеклетки, в антеридиях – сперматозоиды. Для оплодотворения необходима вода. Из зиготы развивается спорофит нового поколения, который первое время питается за счет заростка, потом переходит к самостоятельному питанию. Представители: щитовник мужской; кочедыжник женский; сальвиния; ужовник.

Семенные растения, не образующие цветков и плодов.
К Голосеменным относятся Саговниковые, Гинкговые и Хвойные. Древесные и кустарниковые формы. Листья мелкие (хвоя), часто вечнозеленые. В жизненном цикле доминирует спорофит Семязачатки располагаются на чешуях женских шишек, содержат яйцеклетку и эндосперм (запас питательных веществ – остаток тела женского гаметофита). На чешуях мужских шишек в пыльцевых мешках созревает пыльца. Пыльцевое зерно (мужской гаметофит) имеет воздушные мешки, облегчающие процесс переноса по воздуху. Мужские гаметы (спермии) неподвижны, процесс оплодотворения не зависит от воды. Семена располагаются на чешуях, дополнительных защитных образований не имеют. Произошли от семенных папоротников. Представители: Сосна обыкновенная, ель европейская, лиственница, можжевельник.

Отдел Покрытосеменные (Цветковые).
Семенные растения, имеющие цветки и плоды.
Ныне господствующая группа. Всего более 250 тысяч видов, относящихся к двум классам (Двудольные и Однодольные). Самые разнообразные жизненные формы. В жизненном цикле доминирует спорофит, гаметофит предельно редуцирован (женский представлен зародышевым мешком внутри семязачатка, мужской – пыльцевое зерно). Развивается видоизмененный побег – цветок,- служащий для полового размножения.
Образование пыльцы и зародышевого мешка у большинства растений завершается одновременно. Процессу оплодотворения предшествует процесс опыления. Опыление (перенос пыльцы на рыльце пестика) бывает перекрестное (при помощи ветра или насекомых, иногда при помощи человека) или самоопыление. Для опыления насекомыми характерны яркие крупные цветки с нектаром, крупная липкая пыльца; для опыления ветром характерны мелкие невзрачные цветки, собранные в соцветия, пыльники на длинных тычиночных нитях, перистые рыльца пестиков, огромное количество мелкой легкой пыльцы, цветение до распускания листьев. Попав на рыльце, пыльцевое зерно прорастает в пыльцевую трубку, в которой расположены два неподвижных спермия. Оплодотворение от воды не зависит. На месте цветка созревает плод.

Схема двойного оплодотворения (С.Г. Навашин, 1898 г.)
Источник